




Гликоген способен синтезироваться почти во всех тканях, но наибольшие его запасы находятся в печени и скелетных мышцах.
Накопление данного полисахарида в миоцитах регистрируется в период их восстановления после работы, особенно при приеме богатой углеводами пищи. В гепатоцитах ускорение синтеза гликогена характерно только после приема пищи и при гипергликемии. Такие отличия метаболизма обусловлены наличием изофермента гексокиназы, фосфорилирующей глюкозу в глюкозо-6-фосфат. В печени работает её изоформа – глюкокиназа, обладающая низким сродством к глюкозе, что ведет к захвату моносахарида гепатоцитом только при высокой концентрации в крови (после еды), что впоследствии метаболизирует ее в любом направлении. При нормогликемии преодоление глюкозы цитолеммой клеткой тормозится.
Непосредственно синтез гликогена обеспечивают следующие ферменты.
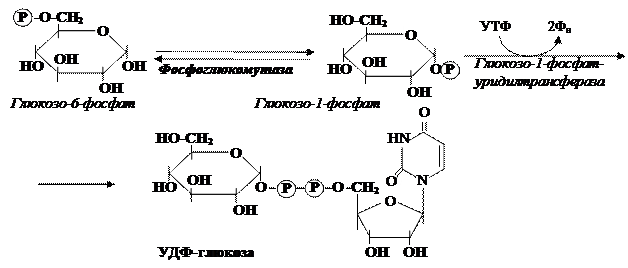
Рис. 9. Реакции синтеза уридилдифосфатглюкозы (УДФ-глюкозы)
Фосфоглюкомутаза обратимо изомеризует глюкозо-6-фосфат в глюкозо-1-фосфат. Глюкозо-1-фосфат-уридилтрансфераза – энзим, осуществляющий ключевую реакцию синтеза. Её необратимость обусловливается гидролизом высвобождающегося при этом дифосфата (рис. 9).
Гликогенсинтаза образует α-1,4-гликозидные связи и удлиняет гликогеновую цепочку, присоединяя первый углеродный атом УДФ-глюкозы к четвертому атому углерода концевого остатка гликогена (рис. 10).
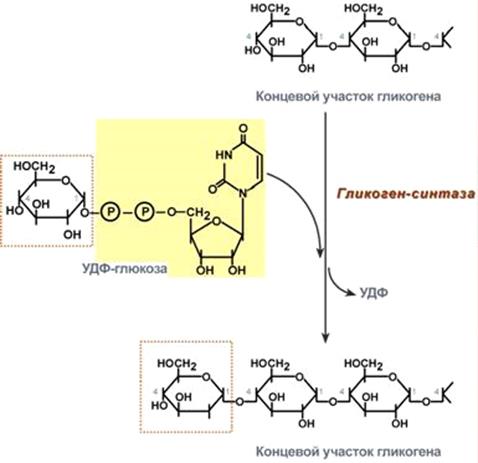
Рис. 10. Химизм реакции гликогенсинтазы
Амило-α-1,4-α-1,6-гликозилтрансфераза (" гликогенветвящий"фермент) переносит фрагмент (6 остатков глюкозы) на соседнюю цепь, образуя α-1,6-гликозидную связь (рис. 11).
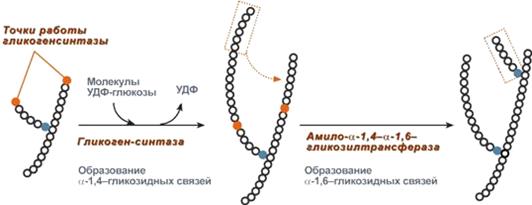
Рис. 11. Роль гликогенсинтазы и гликозилтрансферазы в синтезе гликогена
Гликогенолиз
Гликоген печени расщепляется при снижении концентрации глюкозы в крови, прежде всего между приемами пищи. Через 12-18 часов голодания его запасы в органе полностью истощаются.
В мышцах количество гликогена уменьшается обычно только во время физической нагрузки – длительной и/или напряженной, т.к. этот полисахарид необходим для обеспечения глюкозой работы самих миоцитов. Из-за отсутствия в них глюкозо-6-фосфатазы находящийся в клетках отрицательно заряженный эфир моносахарида не способен преодолеть цитолемму и выйти в кровь, что позволяет использовать гликоген только для собственных нужд.
В гликогенолизе непосредственно участвуют три фермента (рис. 12):
· Фосфорилаза гликогена разрывает α-1,4-гликозидные связи с отщеплением глюкозо-1-фосфата. Фермент работает до тех пор, пока до точки ветвления (α1,6-связи) не останется 4 остатка глюкозы.
· α(1,4)-α(1,6)-Глюкантрансфераза – энзим, переносящий фрагмент трисахарида на другую цепь с образованием новой α1,4-гликозидной связи. При этом на прежнем месте остается один остаток глюкозы и "открытая" действию катализатора доступная α1,6-гликозидная связь.
· Амило-α1,6-глюкозидаза, (" деветвящий " фермент) гидролизует последнюю с отрывом свободной (нефосфорилированной) глюкозы. В результате возникает цепь без ветвлений, вновь служащая субстратом для фосфорилазы.
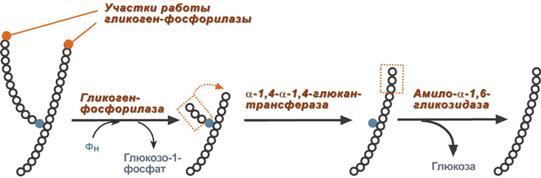
Рис.12. Работа ключевых ферментов гликогенолиза
При этом в одной клетке не могут идти одновременно синтез и распад гликогена – это противоположные процессы с совершенно с разными задачами. Катаболизм и анаболизм гомополисахарида исключают друг друга или, по-другому, они реципрокны.

