




Продукт гликолиза (пировиноградная кислота, или пируват) может использоваться тремя способами.
У аэробных организмов гликолиз составляет лишь первую часть расщепления глюкозы до СО2 и Н2О. В этом случае пируват подвергается окислительному декарбоксилированию — теряет СО2, а двухуглеродный фрагмент (ацетильная группа) включается в ацетилкофермент А и в дальнейшем окисляется в цикле лимонной кислоты до СО2 и Н2О с участием молекулярного кислорода воздуха.
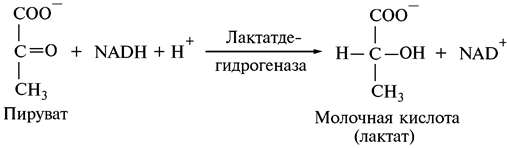
В анаэробных условиях при молочнокислом брожении пировиноградная кислота восстанавливается образовавшимся при гликолизе NADH + H+ (см. реакцию 5), под действием фермента лактатдегидрогеназы получается молочная кислота:
При спиртовом брожении пировиноградная кислота предварительно декарбоксилируется до уксусного альдегида:
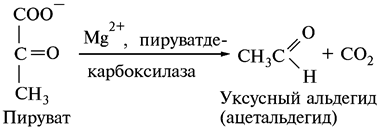
Пируватдекарбоксилаза относится к классу лиаз. Коферментом является тиаминпирофосфат (витамин В1).
Образовавшийся уксусный альдегид восстанавливается до этанола за счет NADH + H+, образовавшегося вследствие окисления глицеральдегид-3-фосфата (см. реакцию 5):
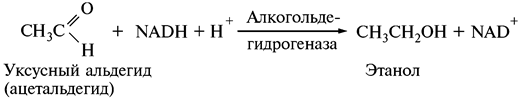
Аэробная фаза дыхания
Аэробный катаболизм глюкозы (дыхание) включает три стадии (рис. 4).
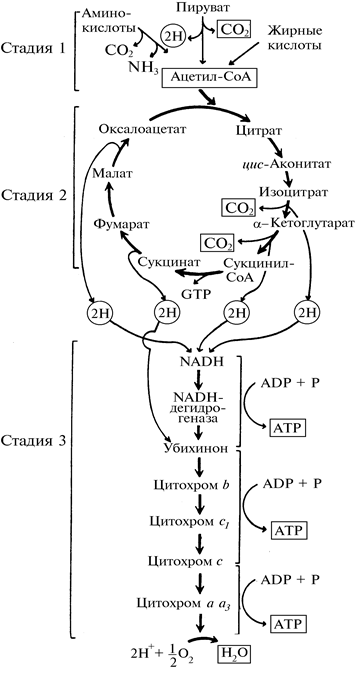
Рис. 4. Три стадии дыхания
На первой стадии из окисляемых молекул глюкозы, жирных кислот и некоторых аминокислот образуются двухуглеродные фрагменты — ацетильные группы, которые затем входят в состав ацетил-кофермента А.
На второй стадии эти ацетильные группы входят в цикл лимонной кислоты, в котором они ферментативным путем расщепляются с образованием высокоэнергетических атомов (протонов и электронов) водорода и высвобождением двух молекул диоксида углерода. Последние представляют конечный продукт окисления органического вещества в процессе дыхания.
На третьей стадии водород в виде протонов и богатых энергией электронов передается по цепи переноса электронов на молекулярный кислород атмосферы. В результате происходит образование АТР (окислительное фосфорилирование), а кислород восстанавливается до воды.
Образование ацетила, активированного коферментом А, из пировиноградной кислоты осуществляется пируватдегидрогеназным комплексом.
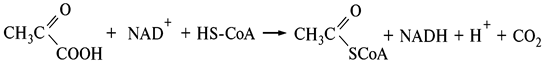
Окислительное декарбоксилирование в физиологических условиях — необратимый процесс.
Ацетил-СоА является также исходным соединением для синтеза жирных кислот и стероидов из углеводов, а также служит ацетилирующим агентом, используемым для разнообразных синтетических процессов в живой клетке.
Цикл лимонной кислоты. Цикл лимонной кислоты, или цикл Кребса (другое название — цикл трикарбоновых кислот), — это циклический процесс (рис. 5). Цикл включает 8 реакций.
1. Конденсация ацетил-СоА с щавелево-уксусной кислотой:
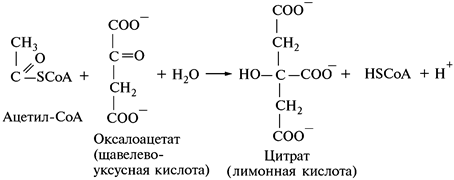
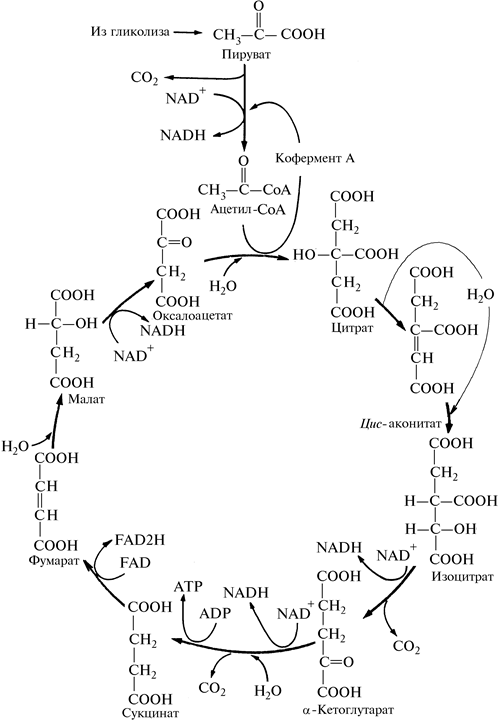
Рис. 5. Цикл Кребса (схема реакций):
В этой реакции метильный углерод ацетильной группы ацетил-СоА связывается с карбоксильной группой щавелево-уксусной кислоты, при этом освобождается кофермент А. Катализатором этой реакции является цитратсинтаза.
2. Превращение лимонной кислоты в изолимонную:
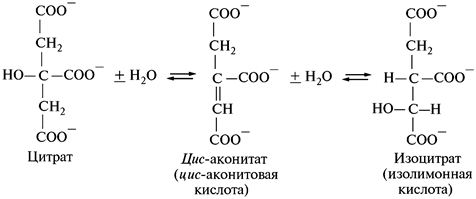
Лимонная кислота превращается в изолимонную через образование цис -аконитовой кислоты под действием фермента аконитазы, которая катализирует обратимое присоединение воды.
3. Окислительное декарбоксилирование изолимонной кислоты:
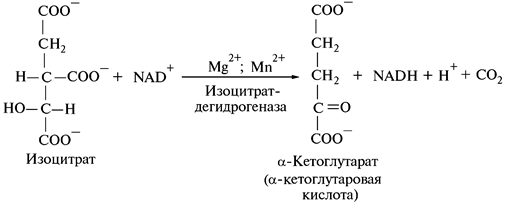
Реакция катализируется изоцитратдегидрогеназой. В качестве промежуточного соединения образуется щавелево-янтарная кислота, прочно связанная с ферментом.
4. Окислительное декарбоксилирование a -кетоглутарата:
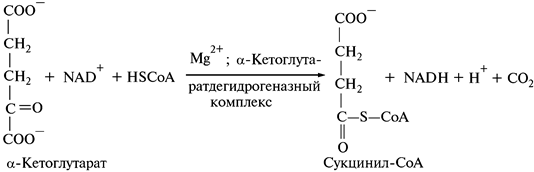
Эта реакция аналогична окислительному декарбоксилированию пировиноградной кислоты. Одновременно с дегидрированием происходит декарбоксилирование a-кетоглутарата. В этой реакции карбоксильная группа, близлежащая к карбонильной, выделяется в виде СО2. Этот углерод принадлежит щавелево-уксусной кислоте.
5. Превращение сукцинил-СоА в сукцинат:
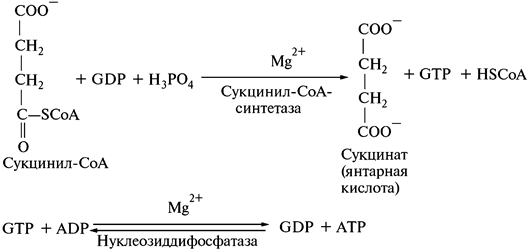
Сукцинил-СоА является высокоэнергетическим соединением. При гидролизе тиоэфирной связи в его составе выделяется энергия, которая накапливается в гуанозинтрифосфате (GTP).
Энергия, содержащаяся в GTP, используется для синтеза АТР.
6. Дегидрирование сукцината:
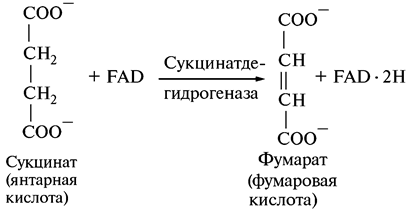
Окисление сукцината катализируется сукцинатдегидрогеназой, единственной реакцией дегидрирования, в которой участвует не NAD+, а FAD.
7. Гидратация фумаровой кислоты:
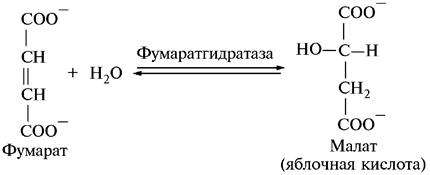
Эта реакция стереоспецифична по отношению присоединения —Н и —ОН воды по двойной связи фумарата и приводит к образованию только L-формы яблочной кислоты.
8. Регенерация щавелево-уксусной кислоты:
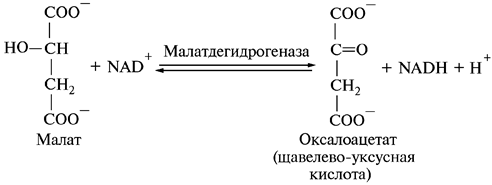
Этой реакцией цикл лимонной кислоты, или цикл Кребса, полностью завершается, и регенерированный оксалоацетат может конденсироваться с новой молекулой ацетил-СоА.
Щавелево-уксусная кислота образуется при карбоксилировании пировиноградной кислоты:

Промежуточные продукты цикла лимонной кислоты используются клеткой в качестве предшественников при синтезе многих биомолекул — аминокислот, жирных кислот, а также терпенов, витаминов и многих других.
Окислительное фосфорилирование. Почти вся энергия, получаемая на ранних этапах окисления субстратов (углеводов или других соединений), вначале запасается в форме высокоэнергетических электронов, переносимых NADH + H+ и FAD·2H. Только позже, в дыхательной цепи, эти электроны взаимодействуют с молекулярным кислородом, высвобождая энергию, используемую ферментами внутренней мембраны митохондрий для синтеза АТР из ADP и неорганического ортофосфата. Эти реакции синтеза АТР проходят в дыхательной цепи и получили название окислительного фосфорилирования.
Энергетика дыхания.
Дыхание — это процесс, в результате которого в живой клетке выделяется свободная энергия.
В живых клетках значительная часть образующейся свободной энергии аккумулируется в высокоэнергетических связях нуклеотидполифосфатов, в частности АТР. АТР затем поставляет энергию для биосинтетических реакций, процессов движения и сокращения.
При дыхании АТР синтезируется как в анаэробной фазе (гликолизе), так и в аэробной.
Синтез АТР из ADP и неорганического фосфора (Н3РО4) в анаэробной фазе называется гликолитическим фосфорилированием, а в аэробной — окислительным фосфорилированием.
Гликолиз является энергетически менее выгодным процессом, чем аэробное дыхание.
В процессах гликолиза выделяется 4 молекулы АТР вследствие того, что глюкоза распадается при гликолизе на две трехуглеродные молекулы — триозы. Однако необходимо учесть, что для активирования гексоз расходуется 2 молекулы АТР. Таким образом, при спиртовом и молочнокислом брожении выделяется всего лишь 2 молекулы АТР (рис. 2,3).
В растениях анаэробные условия встречаются редко, и в клетках высших растений гликолиз протекает нормально в присутствии кислорода. Поэтому NADH + H+, который образовался при окислении глицеральдегид-3-фосфата, не расходуется на восстановление пировиноградной кислоты или уксусного альдегида, а включается в цепь переноса электронов с образованием у растений двух молекул АТР, а в животных организмах — трех.
Из расчета на молекулу глюкозы на этом участке окисления выделяется 4 молекулы АТР. Суммируя молекулы АТР получаем: 2 АТР + 4 АТР = 6 АТР. Это количество свободной энергии, которое получают растительные клетки при окислении глюкозы до пировиноградной кислоты. Основное количество энергии выделяется в аэробных условиях при окислении водорода кислородом воздуха.
При дегидрировании пировиноградной, изолимонной, a-кетоглутаровой, янтарной и яблочной кислот водород отдает электрон и превращается в протон Н+. Электроны водорода по цепи переноса передаются на кислород воздуха, в результате чего образуется молекула воды. Это реакция экзотермическая.
Окисление пировиноградной, изолимонной, a-кетоглутаровой и яблочной кислот происходит под действием дегидрогеназ, коферментом которых является NAD+, при этом NAD+ восстанавливается до NADH + H+ (длинная дыхательная цепь).
Энергия, высвобождающаяся при переносе одной пары электронов отNADH + H+ к кислороду, запасается в виде трех молекул АТР. Таким образом, окисление пировиноградной, изолимонной, a-кетоглутаровой и яблочной кислот сопровождается образованием 12 молекул АТР.
В окислении янтарной кислоты NAD+ участия не принимает. Оно начинается с NADH-зависимой дегидрогеназы, коферментом которой является FAD (короткая дыхательная цепь). Поэтому в данном случае образуется только 2 молекулы АТР. Кроме того, в цикле лимонной кислоты, или цикле Кребса, выделяется 1 молекула АТР на уровне субстратного фосфорилирования при образовании янтарной кислоты из сукцинил СоА. Следовательно, при полном окислении одной молекулы пировиноградной кислоты получается 15 молекул АТР.
Суммируя молекулы АТР, образовавшиеся при окислении глюкозы до пирувата (6 молекул АТР) и при окислении пирувата до СО2 и Н2О (30 молекул АТР), получаем 36 молекул АТР. С учетом того что при гидролизе АТР до ADP выделяется 30,5 кДж, эффективность запасания свободной энергии при аэробном дыхании за счет глюкозы составляет 38,3 %.
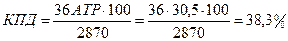
Энергетический выход при расщеплении одной молекулы глюкозы:
| Гликолиз | 2 АТР |
| 2NADH+H+ ® 4 ATP ® 6 ATP | |
| Пируват ® Ацетил-СоА | 1NADH+H+ ® 3 ATP (х2) ® 6 ATP |
| Цикл Кребса | 1 АТР |
| 3NADH+H+ ® 9 ATP (х2) ® 24ATP | |
| FAD·H ® 2 ATP |

