




Транспортные РНК.
Каждая клетка содержит до 60 видов молекул тРНК. Они самые маленькие из всех видов НК (М = 25000), содержит 73 – 93 нуклеотида. Функции тРНК заключаются в транспортировке аминокислот к месту синтеза белка – в рибосомы. тРНК обладают специфичностью. Аминокислота, транспортируемая тРНК доставляется к определенному участку информационной (матричной РНК), указанному кодоном. Чтение (распознавание) кодонов достигается вследствии наличия в каждой тРНК уникальной тринуклеотидной последовательности, называемой антикодонами, последовательность которого комплементарна последовательности кодона. Пространственная структура тРНК напоминает «клеверный лист»:
 АК
АК
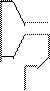 | 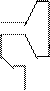 |


 антикод
антикод
тРНК содержит до 50 различных нетипичных (модифицированных) нуклеотидов. Присоединение АК к тРНК катализируется ферментом аминоацил-тРНК-синтетазой. И уже в виде комплекса аминоацил-тРНК аминокислота доставляется к месту синтеза белка.
(3) Рибосомные РНК. По сути, рибосоы – мультимолекулярные агрегаты, состоящие из белка (35%) и рРНК (65%). рРНК составляет до 80% от всех РНК клетки. Диаметр рибосом достигает до 50 Ǻ. Для всех живых организмов характерны два вида рибосом: 70S (у микробов) и 80S (у других видов). Интактная рибосома представляет собой комплекс из двух субъединиц – тяжелой (50S) и легкой (30S). Весь этот интактный комплекс может диссоциировать на субъединицы, которые в свою очередь могут диссоциировать с образованием до 60 компонентов (это молекулы РНК и различных белков). Эти компоненты способны реконструироваться в субъединицы, которые далее участвуют в реконструкции рибосом. Это естественный процесс происходящий в клетках, причем протекающий самопроизвольно.
Структура ДНК.
ДНК предназначена для хранения и передачи генетической информации. По химической структуре ДНК, в отличие от РНК содержит дезоксирибозу, и вместо урацила – тимин. М (ДНК) велика и составляет от 10 в 6 до 10 в 9 степени и более. Размеры молекулы также очень вариабельны и среди различных видов и внутри одного вида. Так, длина ДНК кишечной палочки составляет от 1 до 13 мм, у человека может достигать 8 см если их вытянуть в длину, а диаметр равен 20 Ǻ, то-есть достаточно чтобы увидеть в электронный микроскоп. Для нуклеотидного состава ДНК существуют общие закономерности, известные как правила Чаргаффа:
1) число пуриновых оснований (А+Г) = числу пиримидиновых (Ц+Т), то есть
(А+Г)/(Ц+Т) = 1
2) число остатков аденина равно числу остатков тимина, то есть А/Т=1.
3) Число остатков гуанина равно числу остатков цитозина, то есть Г/Ц=1.
В 1953 г. Уотсон и Крик показали, что молекула ДНК состоит из двух правозакрученных спиральных полинуклеотидных цепей, переплетенных друг с другом и противоположно направленых. Эта структура двойной спирали стабилизирована межцепочечными водородными связями между спецефическими парами АО, которые направлены от сахарофосфатного остова каждой цепи внутрь спирали перперндикулярно оси. Это полностью соответствует правилу Чаргоффа. Аденин всегда связан двумя водородными связями с тимином А=Т (поэтому и соотношение А/Т=1), а гуанин всегда связан тремя водородными связями с цитозином Г º Ц (поэтому соотношение Г/Ц=1). В соответствии с этим эти пары называются комплементарными парами оснований. Более того, полная последовательность одной из цепей полностью комплементарна последовательности второй. Две цепи имеют противоположную направленность – одна из них ориетирована в 3` - 5` -направлении, а вторая – в 5` - 3` -направлении. Важно, что именно противоположная направленность двух цепей обеспечивает правильную пространственную ориентировку АО. Водородные связи – не единственный вид взаимодействия, стабилизирующего двухцепочечную структуру. Молекула ДНК – полианион, на ее поверхности локализовано множество отрицательных зарядов, что обеспечивает стабилизацию путем электростатических взаимодейтвый с неорганическими поликатионами (например с ионами магния) или с белками (гистонами), содержащими большое количество положительно заряженных боковых цепей АК. Третий стабилизирующий фактор возникает благодаря гидрофобным взаимодействиям в плотно упакованных АО, которые уложены как бы стопкой в сердцевине спирали. На один виток двойной спирали приходится 10 нуклеотидов. Двойная спираль закольцована. Исследования последних лет показали, что двойная спираль не только представляет собой кольцевую структуру, но и это кольцо тоже закручено (в этом случае говорят о суперскрученной или суперспирализованной ДНК). Один полный виток «суперзакрутки приходится на 20 – 25 витков двойной спирали, то есть в одном витке суперспирали 200 – 250 пар АО. Суперскрученная ДНК имеет существенное значение для многих процессов. Кроме того, суперспирализация объясняет, каким образом массивная молекула ДНК способна упаковаться в маленьком объеме бактериальной клетки или ядре. В то же время, существует и достаточно основательные предположения, что нативная ДНК может иметь до 10 других различных форм (может быть левозакрученной, на один виток может приходится разное количество АО и т.д.). Для ДНК характерно явление денатурации: при различных химических или физических воздействиях двухцепочечная ДНК может быть раскручена. Денатурирующие агенты те же, что и факторы вызывающие денатурацию белков, то есть ионизирующее излучение, ультразвук, соли тяжелых металлов, концентрированные кислоты и щелочи и т.д. При этом двойная спираль раскручивается и цепи расходятся. Денатурация может быть обратимой (если убрать денатурирующий агент, то структура ДНК восстанавливается). При этом возникает явление ренатурации, то есть разошедшиеся цепи денатурированной ДНК снова объединяются в двойные спирали не отличающиеся от исходных. Но при этом возможна гибридизация ДНК. То есть можно получить такие молекулы ДНК, которые образованы двумя цепями, первоначально принадлежавшие молекулами ДНК разных видов, при условии, что эти ДНК близки друг к другу и их комплиментарные цепи обладают достаточной аналогией для того чтобы обеспечить их попарное соединение. Возможно и ДНК – РНК - гибридизация так же при условии достаточной комплиментарности.
Распад НК.
1) Переваривание. НК поступают в организм, как правило, в виде нуклеопротеидов с пищей. Под влиянием протеаз желудка и кишечника, с участием соляной кислоты, нуклеопротеиды пищи распадаются на белок и НК. Белковая часть подвеоргается гидролитическму расщеплению до свободных АК, аналогично другим белкам. Распад же НК происходит, в основном, гидролитическим путем в тонком кишечнике под действием ферментов нуклеаз. Различают рибонуклеазы (РНК - азы) – они специфичны только по отношению к РНК, другие – дезоксорибонуклеазы (ДНК - азы) – действуют только на ДНК. Третья группа неспецифических нуклеаз активна по отношению к обоим видам НК. В распаде ДНК участвуют различные виды ДНК – аз; одни из них относятся к эндонуклеазам, другие – к экзонуклеазам. Так, из поджелудочной железы выделена ДНК – аза – 1, она активна в нейтральной среде и разрывает цепь молекулы ДНК преимущественно между пуриновыми и пиримидиновыми нуклеотидами. Разрыв происходит между фосфатом и 3` - гидроксилом дезоксирибозы. ДНК – аза –2 получена из селезенки и тимуса. Оптимум рН лежит в кислой среде, разрывает цепь молекулы ДНК между фосфатом и 5`-гидроксилом дезоксирибозы. Локализована в лизосомах. И ДНК-аза-1 и ДНК-аза-2 являются эндонуклеазами, для их работы необходимо присутствие ионов магния или марганца и после их действия образуются ди и олигонуклеотиды. Последующее расщипление осуществляется экзонуклеазами или, как их еще называют, фосфодиэстеразами. Они последовательно отделяют нуклеотиды с одного из концов молекулы ДНК, образуя мононуклеотиды.
РНК-аза разрывает цепь РНК между фосфатом и 5` гидроксилом рибозы. РНК-аза также относится к эндонуклеазам. В результате ее действия образуются олигонуклеотиды. В дальнейшем олигонуклеотиды подвергаются действию экзонуклеаз и расщепляются до мононуклеотидов. Таким образом в результате действия эндо- и экзонуклеаз молекулы ДНК и РНК расщепляются до моно нуклеотидов. Освобожденные нуклеотиды гидролизуются кишечными кислотными и основными фосфатазами или нуклеотидазами до нуклеозидов и фосфата. Нуклеозиды и фосфаты всасываются слизистой кишечника, попадают с током крови в ткани и там, если нуклеозиды не используются для синтеза НК, подвергаются действию нуклеозидаз, расщепляющим связь между АО и углеводным компонентом. В клетках распад идет аналогично. Таким образом в результате переваривания нуклеиновых кислот или их внутриклеточного распада получаются АО, фосфат и рибоза, либо дезоксирибоза. Фосфат используется для различного рода реакций фосфорилирования, углеводы используются в углеводном обмене, поступают, например, в пентозо-фосфаный путь обмена углеводов или идут на синтез новых НК. АО также могут использоваться для синтеза новых НК, однако, чаще всего они распадаются, а для синтеза НК используются АО, синтезированные из низкомолекулярных азотистых и безазотистых предшественников.
2) Распад пуриновых оснований.
NH2 OH
 | |
| |




 N N + H2O; NH3 N N +O2; + H2O; -H2O2
N N + H2O; NH3 N N +O2; + H2O; -H2O2
 |  |
адениндезаминаза ксантиноксидаза
N NH (у бактерий) N NH
Аденин гипоксантин
 NH2 OH
NH2 OH
 | |
| |
N N + H2O; NH3 N N +O2; + H2O; -H2O2
| |
аденозиндезаминаза нуклеозидаза

 N N (у человека) N N
N N (у человека) N N


 НОН2С О НОН2С О
НОН2С О НОН2С О

 Н Н Н Н Н Н Н Н
Н Н Н Н Н Н Н Н
НО ОН НО ОН
Аденозин инозин





 ОН
ОН
 | HOH2C O OH
| HOH2C O OH
 N N
N N
+ H H H
N OH OH
NH
гипоксантин рибоза

ОН OH
| |
N N +O2; + H2O; -H2O2 N N
 Ксантиноксидаза
Ксантиноксидаза
 HO HO
HO HO
НО N NH N NH
Ксантин Мочевая кислота
ОН OH
| |
 N N +H2O; -NH3 N N и далее
N N +H2O; -NH3 N N и далее
 гуаназа
гуаназа
HO
NH2 N NH N NH
Гуанин Ксантин
В тканях человека найдена аденозинаминаза, в этом случае происходит дезаминирование аденозина с образованием инозина, а дальше действует нуклеозидаза расщепляя инозин до гипоксантина и рибозы.
У человека мочевая кислота является конечным продуктом распада пуриновых оснований. У большинства наземных животных мочевая кислота окисляется в аллантоин, у рыб далее окисление идет до аллантоиновой кислоты, а в микроорганизмах – до мочевины. В норме в сыворотке крови содержится мочевой кислоты: у мужчин 0,24 – 0,50 ммоль/л, у женщин – 0,15 – 0,40 ммоль/л. Растворимость мочевой кислоты примерно 0,60 ммоль/л. Экскреция мочевой кислоты за сутки составляет 0,35 – 1,5 г. Мы уже упоминали о том, что часть азотистых оснований не распадается, а реутилизируется, то есть вновь используется на синтез нуклеотидов. Выделяют два основных фермента реутилизации пуриновых оснований:
Аденинфосфорибозилтрансфераза и гипоксантин-гуанинфосфорибозилтрансфераза. Оказалось, что врожденный генетический дефект – отсутствие второго фермента (гипоксантин-гуанинфосфорибозилтрансфераза) приводит к повышенному синтезу мочевой кислоты, проявляющемуся клинически как синдром Леш-Нихана, характеризующийся повышенной нервозностью, агрессивностью, замедлением умственного развития. В результате повышенного синтеза мочевой кислоты и неспособностью почек выводить ее, происходит накопление и кристаллизация мочевой кислоты в синовиальной жидкости вокруг суставов (особенно большого пальца ноги), вызывая боль, тугоподвижность сустава, (подагра) и т.д. Накопление мочевой кислоты в почечных канальцах может привести к развитию мочекаменной болезни. Для лечения применяют аллопуринол, снижающий синтез мочевой кислоты путем конкурентного ингибирования ксантиноксидазы. Повышение содержания мочевой кислоты отмечают и при других патологиях: тяжелые инфекционные заболевания, поражения почек и т.д. Повышается содержание мочевой кислоты в крови и в период новорожденности. Снижение содержания мочевой кислоты отмечено при акромегалии, болезни Вильсона, при инъекциях инсулина и т.д.
4) Распад пиримидиновых оснований:
NH2 O O

 | || НАДФН2 НАДФ ||
| || НАДФН2 НАДФ ||


 N +Н2О; -NH3 HN CH HN CH +Н2О
N +Н2О; -NH3 HN CH HN CH +Н2О
 |  | |
O= цитозин- O= CH дегидро- O= CH2 гидролаза
NH дезаминаза NH урацил(ДГ) NH
цитозин урацил дгидроурацил
NH2
|
C=O
|
NH +H2O
 | NH3 + CO2 + b-аланин.
| NH3 + CO2 + b-аланин.
CH2 гидролаза
|
CH2
|
COOH
Карбомоил-b-аланин.
b-аланин либо подвергается дальнейшему распаду с образованием аммиака, углекислого газа, воды, либо используется на синтез КоА, карнозина и т.д. Тимин распадается аналогично, но при распаде дает не b-аланин, а b-аминомасляную кислоту: NH2 – CH2 – CH – COOH
|
CH3
Поскольку из аммиака, образовавшегося при распаде пиримидиновых оснований, синтезируется мочевина, то ее и можно считать конечным продуктом распада пиримидиновых оснований. В норме содержание мочевины в сыворотки крови составляет 2,5 – 8,3 ммоль/л. Суточная экскреция с мочой: 20 – 35 г. Поскольку конечным продуктом распада пиримидиновых оснований (NH3, CO2, бета-аланин) хорошо растворимы в воде, то клинические симптомы, связанные с нарушением распада пиримидинов выражены слабо. Повышение содержания мочевины в сыворотке отмечают при гемолитической анемии, отравлении и т.д. Снижение содержания мочевины в сыворотке отмечают при генетических нарушениях цикла образования мочевины. Повышение содержания мочевины в моче связаны с гипертиреозом, постоперационными состояниями. Снижается мочевина в моче при гепатопатиях, почечной недостаточности, лечении инсулином.
СИНТЕЗ НУКЛЕИНОВЫХ КИСЛОТ.
1) Синтез пиримидиновых нуклеотидов. Для того, чтобы синтезировать нуклеиновую кислоту, представляющую собой, как известно, полинуклеотидную цепочку, необходимо синтезировать отдельные нуклеотиды. Для синтеза пуриновых и пиримидиновых нуклеотидов могут использоваться либо уже готовые азотистые основания, углеводы и фосфорная кислота, поступающие, например, в клетку из ЖКТ, или образующиеся при внутриклеточном распаде НК, либо для синтеза используются простые соединения: СО2, аммиак, аспартат, глицин, глютамин, рибоза. Существует два отдельных пути синтеза нуклеотидов – один для пиримидинов, другой для пуринов. Но если для синтеза пуринов в клетке есть ферменты, способные утилизировать для синтеза нуклеотидов готовые пуриновые основания, то клетки не располагают эффективными путями для использования свободных пиримидиновых оснований для синтеза нуклеотидов. В то же время они обладают способностью использовать пиримидиновые рибонуклеозиды уридин и цитидин. При этом достаточно добавить фосфорную кислоту и нуклеотид готов. Этот процесс каталиируют соответствующие ферменты: уридинкиназа и цитидинкиназа. Однако гораздо чаще пиримидиновые АО синтезируются из «предшественников». Первым продуктом нуклеотидной природы пиримидинового пути является уридин – 5 – монофосфат, которые затем служит предшественником всех остальных пиримидиновых нуклеотидов. Фосфорибозный компонент для синтеза уридин – 5 – монофосфата переносится в виде 5 – фосфорибозил – 1 – пирофосфат, который, в свою очередь, получается из рибозо – 5 – фосфата, а рибоза – 5 – фосфат из глюкозы, скажем, через пентозофосфатный путь. Схематически этот процесс можно представить так:


 Гексозы……. НО3РОСН2 О ОН АТФ АМФ
Гексозы……. НО3РОСН2 О ОН АТФ АМФ

Н Н Н Н фосфорибозил
трансфераза
ОН ОН
Рибоза – 5 - фосфат

 Н2О3РОСН2 О
Н2О3РОСН2 О
 О О О
О О О
Н Н || ||
Н Н Р – О – Р – ОН
| |
ОН ОН OH OH
5 – фосфорибозил – 1 – пирофосфат
Для дальнейшего синтеза пиримидиновых нуклеотидов необходимо синтезировать циклическую пиримидиновую структуру. Первым этапом синтеза является образования карбомоилфосфат:

 2АТФ 2АДФ –H3PO4 NH2
2АТФ 2АДФ –H3PO4 NH2

 NH3 + CO2 C OPO3H2
NH3 + CO2 C OPO3H2
 карбомоилфосфатсинтетаза ||
карбомоилфосфатсинтетаза ||
из глютамина O
С этим продуктом (карбомоилфосфат) вы уже встречались в ходе орнитинового цикла, как с промежуточным продуктом в процессе синтеза мочевины. Однако, карбомоилфосфат, необходимый для синтеза мочевины образуется в митохондриях при участии митохондриального фермента карбомоилфосфатсинтетазы – 1, а карбомоилфосфат, участвующий в биосинтезе пиримидиновых оснований, образуется в цитозоле под действием другой формы того же фермента – карбомоилфосфатсинтетазы – 2. Образовавшийся в цитозоле карбомоилфосфат взаимодействует с аспартатом, образуя N – карбомоиласпартат
NH2 COOH NH2 COOH
| | | |

 C = O + CH2 -H3PO3 C CH2
C = O + CH2 -H3PO3 C CH2
| | || |
 OPO3H2 H2N–CH аспартаттранс O HN CH
OPO3H2 H2N–CH аспартаттранс O HN CH
Карбомоил аспартат карбомоилаза |
фосфат COOH
N – карбомоиласпартат
Далее происходит дегидротация и циклизация:
NH2 COOH O O
| | || ||


 C CH2 - H2O C НАДН2 C
C CH2 - H2O C НАДН2 C

 || HN | NH CH2 НАД HN CH
|| HN | NH CH2 НАД HN CH
 O CH дигидроортаза | | | ||
O CH дигидроортаза | | | ||
 | C CH дигидрооротат C C-COOH
| C CH дигидрооротат C C-COOH
 COOH || NH ДГ || NH
COOH || NH ДГ || NH
O COOH O
Дигидрооротовая кслота оротовая кислота
На этой стадии к оротовой кислоте присоединяется уже полученный нами ранее
5 – фосфорибозил – 1 – пирофосфат, с образованием оротидиловой кислоты:
O
||

 C Н2О3Р– СН2 О О O O
C Н2О3Р– СН2 О О O O
HN CH || || - ПФ
 | || + Н Н Н Н Р – О – Р - ОН
| || + Н Н Н Н Р – О – Р - ОН

 C C-COOH | | оротофосфорибозил-
C C-COOH | | оротофосфорибозил-
|| NH OH OH трансфераза
O ОН ОН
оротовая кислота 5 – фосфорибозил – 1 – пирофосфат
O O
|| ||
C C
NH CH -СО2 HN CH
 | || | ||
| || | ||
 C C оротидилат C CН
C C оротидилат C CН

 || NH декарбоксилаза || NH
|| NH декарбоксилаза || NH
O COOH O
Н2О3РН2С О Н2О3РН2С О
| | | |
 Н Н Н Н
Н Н Н Н
Н Н Н Н
ОН ОН ОН ОН
Оротидиловая кислота Урацил – 5 – монофосфат (УМФ)
Необходимо отметить, что из 6 ферментов, участвующих в синтезе УМФ, первые 3 (карбомоилфосфатсинтетаза, аспартаттранскарбомоилаза и дегидрооротаза) связаны в составе одной полипетидной цепи. Два же последних фермента (оротатфосфорибозилтрансфераза и оротидилатдекарбоксилаза) образуют другой комплекс. Суммарно же процесс синтеза УМФ можно представить:

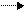 CO2 + NH3 + АСПАРТАТ 2АТФ 2АДФ O
CO2 + NH3 + АСПАРТАТ 2АТФ 2АДФ O
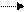
 +5 – фосфорибозил- || из аспартата
+5 – фосфорибозил- || из аспартата
 - 1 – пирофосфат 6 ферментов C
- 1 – пирофосфат 6 ферментов C
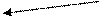 HN CH
HN CH
 Из NH3 | || УМФ
Из NH3 | || УМФ
 C CH
C CH
 || N
|| N
из СО2 O |
Рибоза – Н3РО4
из ФРПР
Но для синтеза НК необходимы трифосфаты. Уридинтрифосфат полуается из УМФ путем нуклеозидкиназных реакций:
 АТФ АДФ АТФ АДФ
АТФ АДФ АТФ АДФ
 УМФ УДФ УТФ
УМФ УДФ УТФ
нуклеозидкиназа нуклеозидкиназа
 Так же, нукледифосфаты и нуклеозидтрифосфаты могут взаимно превращаться под действием нуклеозид-дифосфат-киназ: УДФ + АТФ УТФ + АДФ.
Так же, нукледифосфаты и нуклеозидтрифосфаты могут взаимно превращаться под действием нуклеозид-дифосфат-киназ: УДФ + АТФ УТФ + АДФ.
нуклеозид-дифосфат-киназ
Синтез ЦТФ. Из УТФ образуется ЦТФ путем аминирования в положении 4 пиримидинового кольца. Источником азота может быть глютамин (превращаясь при этом в глютомат) или же просто аммиак. Схематично этот процесс выглядит так:
O NH2
|| |
C C
 NH CH АТФ АДФ + Фк HN CH
NH CH АТФ АДФ + Фк HN CH
| || | ||
C C +NH3; -H2O C CН

 || NH (глютамин) || NH
|| NH (глютамин) || NH
O O
рибоза глютамат рибоза
| |
Ф – Ф – Ф ЦТФ - синтетаза Ф – Ф – Ф
УТФ ЦТФ.
Синтез ТТФ.
Известно, что тиминовые нуклеотиды содержатся в ДНК, то есть в их состав входит не рибоза, а дезоксирибоза. Соответственно, синтезируются ТМФ из нуклеотиддифосфатов, содержащих не рибозу, а дезоксирибозу. Мы уже говорили, что предшественником всех пиримидиновых нуклеотидов является уридин – 5 –монофосфат. Путем нуклеозидкиназной реакции, как было показано выше, УМФ превращаются в УДФ. Но для того чтобы УДФ превратить в дУДФ, то есть нуклеотиддифосфат, содержащий дезоксирибозу, необходимо участие трех белков: тиоредоксинредуктазы, тиоредоксина и рибонуклеотидредуктазы. Схемы действия этих белков относительно проста: тиоредоксинредуктаза катализирует восстановление тиоредоксина, тиоредоксин (восстановленный) активирует рибонуклеотидредуктазу, а она, в свою очередь, способствует восстановлению рибозы (в составе нуклеотиддифосфата) до дезоксирибозы. Таким образом, мы получаем дУДФ. дУДФ переходит в дУМФ, который подвергается метилированию. Донором метильной группы является N(5), N(10) – метилен – FН4 (тетрогидрофолвая кислота).
O О
| | ||||||
| | ||||||
 НN N(5), N(10)-CH2 – FH4 НN СН3
НN N(5), N(10)-CH2 – FH4 НN СН3
| |||
 | |||

O N тимилатсинтетаза O N
 дезоксирибозы. дезоксирибозы.
дезоксирибозы. дезоксирибозы.
Н3РО4 Н3РО4
дЦМФ дТМФ
В дальнейшем для получения ТДФ и ТТФ можно привести две стадии фофорилилрования (с использованием нуклеозидкиназ). Получающиеся в результате реакции образования ТМФ гидрофолат (FH2) необходимо восстановить снова до тетрагидрофолата (FH4). Это осуществляется под действием фолатредуктазы и НАДФН. На ингибировании фолатредуктазы (а, соответственно, и образовании ТМФ) основано действие группы антифолатных средств, оказавшихся эффективными противоопухолевыми препаратами. Особенно эффективен: аминоптерин и метотрексат. Тимидилатсинтетаза (фермент, катализирующий реакцию
 дУМФ дТМФ) непосредственно ингибируется 5-фторурацилом и 5-фторо-2`- дезоксиуридином, что тоже используется в противораковой терапии. Таким образом недостаток тимидина и FH4 в быстрорастущих и делящихся раковых клетках останавливает клеточный рост и даже приводит к гибели клеток. Биосинтез пиримидиновых нуклеотидов регулируется ингибированием по принципу обратной связи. Схема регуляции:
дУМФ дТМФ) непосредственно ингибируется 5-фторурацилом и 5-фторо-2`- дезоксиуридином, что тоже используется в противораковой терапии. Таким образом недостаток тимидина и FH4 в быстрорастущих и делящихся раковых клетках останавливает клеточный рост и даже приводит к гибели клеток. Биосинтез пиримидиновых нуклеотидов регулируется ингибированием по принципу обратной связи. Схема регуляции:
ингибитор ингибитор
ЦМФ ЦТФ

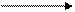
 СО2 + глютамин + АТФ карбомоил- N-карбомоиласпартат
СО2 + глютамин + АТФ карбомоил- N-карбомоиласпартат
(NН3) карбомоил- фосфат аспартат-
фосфат-синтаза транскарбомоилаза (АТК-аза)
 УМФ УТФ ЦТФ
УМФ УТФ ЦТФ
ЦТФ ингбирует АТК-азу, снижая сродство этого фермента к субстрату. АТФ, напротив, активирует АТК-азу, повышая сродство этого фермента к субстрату. При этом идет конкуренция АТФ и ЦТФ за активный центр фермента. АТФ в высокой концентрации вытесняет ЦТФ. Это имеет высокий биологический смысл: (1) активация АТК-азы под действием АТФ уравнивает скорости образования пуриновых и пиримидиновых нуклеотидов (а для синтеза НК нужны их сопоставимые количества); (2) активация под действием АТФ сигнализирует о его досточной концентрации в качестве субстрата для некоторых реакций биосинтеза пиримидиновых нуклеотидов (например, синтеза карбомоилфосфата, фосфорилирования УМФ до УТФ).
2) Синтез пуриновых нуклеотидов. Мы уже упоминали, о том, что для синтеза пуриновых нуклеотидов могут использоваться готовые пуриновые азотистые основания, либо пуриновые нуклеозиды, либо идет синтез из предшественников. В первом случае используются два фермента: а) аденинфосфорибозилтрансфераза (переносит фосфорибозу с ФРПФ на аденин с образованием АМФ). б) гипоксантин-гуанинфосфорибозилтрансфераза (переносит фосфорибозил с ФРПФ на гипоксантин и гуанин с образованием инозинмонофосфата и ГМФ соответственно).
Причем второй фермент работает более активно. А при генетическом дефекте (его отсутствии) развивается синдром Леш-Нихана, возможно развитие мочекаменной болезни, подагры. Если же для синтеза пуриновых нуклеотидов используется готовый нуклеозид, то, как и в случае пиримидиновых нуклеотидов работают киназы (например, аденозинкиназа), катализирующие присоединение к нуклеозиду остатка фосфорной кислоты. Процесс биосинтеза пуриновых оснований из предшественников несколько отличается от синтеза примидиновых оснований. Если, в случае синтеза пиримидиновых нуклеотидов, 5-фосфорибозил – 1- пирофосфат присоединялся на одной из последних стадий образования уридин – 5`-монофосфата – первого продукта нуклеотидной природы, образующегося в ходе пиримидинового пути, то при синтезе первого продукта нуклеотидной природы пуринового пути – инозин - 5`-монофосфата (УМФ) – 5 – фосфорибозил – 1 – пирофосфат принимает участие уже в первой реакции:
Н2О3РОСН2 О Н2О3РОСН2 NH2
 О О О О
О О О О
Н Н || || (1) Н Н
 Н Н Р – О – Р – ОН (2) H H
Н Н Р – О – Р – ОН (2) H H
| |
ОН ОН OH OH OH OH
5 – фосфорибозил – 1 – пирофосфат 5-фосфорибозил – 1 – амин
(1) – +глютамин; -глютамат; - ПФ
(2) – амидофосфорибозилтрансфераза.
+Затем 5-фосфорибозил – 1 – амин вступает в реакцию с глицином.
Н2О3РОСН2 NH3 (+) Н2О3РОСН2 NH–C–CH2
 О АТФ АДФ+Фк || |
О АТФ АДФ+Фк || |
 Н Н +глицин –Н2О Н Н O NH2
Н Н +глицин –Н2О Н Н O NH2
Н Н H H
ОН ОН OH OH
5-фосфорибозил – 1 – амин рибонуклеотид
глицинамида
(глицинамидрибозилфосфат)
 … УМФ
… УМФ
Далее идет еще 7 реакций до образования инозин – 5 – монофосфата:
СО2, аспартат, глицин, 2 глютамина, 2 тетрогидрофолата
5 – фосфорибозил –1 – пирофосфат
 |
4 АТФ
 |
10 ферментов
4 АДФ
+ Из СО2
 4 Н3РО4
4 Н3РО4
 О Из глицина
О Из глицина



 Из аспартата ||
Из аспартата ||
 HN N
HN N



 Из FH4 Из FH4
Из FH4 Из FH4


 N N
N N
 |
|
 Рибоза - ФК
Рибоза - ФК
Из глютамина Из 5 – фосфорибозил – 1 – пирофосфата
Далее, из инозин –5 – фосфата возможен синтез АМФ и ГМФ:
O O NH
 || +НАД НАДН2 || - H2O |
|| +НАД НАДН2 || - H2O |

 HN N +НО2 HN N +ионMg HN N
HN N +НО2 HN N +ионMg HN N
 ГТФ ГДФ + Н3РО4
ГТФ ГДФ + Н3РО4
 ДГ Аспартат
ДГ Аспартат
|| (УМФ - ДГ)
 O NH N N N Фумарат N N
O NH N N N Фумарат N N
Рибоза Рибоза Рибоза
| | |
ФК ФК ФК
(Ксантозин-5-монофосфат) (Инозин-5-монофосфат) (АМФ)

 2АТФ Глютамин АТФ Нуклеотид-
2АТФ Глютамин АТФ Нуклеотид-

 киназа
киназа
2АДФ
+ Глютамат АДФ
 2Н3РО3 (АДФ)
2Н3РО3 (АДФ)
О АТФ Нуклеотид-
 || Киназа
|| Киназа
HN N
 АДФ
АДФ
 (АТФ)
(АТФ)
H2N
 N N
N N
Рибоза
|
 ФК АТФ АДФ АТФ АДФ
ФК АТФ АДФ АТФ АДФ
 (ГМФ) Нуклеотидкиназа (ГДФ) Нуклеотидкиназа (ГТФ)
(ГМФ) Нуклеотидкиназа (ГДФ) Нуклеотидкиназа (ГТФ)
Образование дезокси – пуриновых нуклеотидов идет так же, как и образование дезокси – пиримидиновых нуклеотидов: на уровне ди- фосфатов участвуют 3 белковых фактора (рибонуклеотидредуктаза – активируется восстановленной формой тиоредоксина, который в свою очередь восстанавливается с помощью тиоредоксинредуктазы). Биосинтез пуриновых нуклеотидов тоже регулируется по типу обратной связи (как и биосинтез пиримидиновых нуклеотидов). Так, накопление АМФ и ГМФ, действует ингибирующе на амидофосфорибозилтрансферазу, фермент, который катализирует первую реакцию синтеза пуриновых оснований (реакцию переноса аминогруппы с глютамина на 5-фосфорибозил – 1- пирофосфат). Второй регуляторный механизм действует на одной из более поздних стадий синтеза пуриновых нуклеотидов. Так, избыток ГМФ в клетке вызывает аллостерическое ингибирование процесса образования ГМФ из инозиновой кислоты, но при этом не влияет на синтез АМФ из инозин – 5 – монофосфата. Избыток же АМФ ингибирует синтез АМФ из инозин – 5 – монофосфата, но не влияет на синтез ГМФ.

