




Лекция 2
Роль воды в жизнедеятельности растений
Гомеостатический порог оводненности растений (ниже которого организм гибнет) для всех растений достаточно высок и составляет для гигрофитов 65-70%, для мезофитов -45-6-%, ксерофитов – 25-27%. В норме вода составляет 70-99% сырой массы растительных тканей (в мезофите листьев до 85, в корне до 99%), выполняя первостепенную роль во всех процесса жизнедеятельности:
А) объединяет все части организма, начиная от молекул в клетках и до раздела жидкость-газ в листьях, где она испаряется;
Б) является важнейшим растворителем и средой для протекания биохимических реакций;
В) участвует в упорядочении структур в клетках. Она входит в состав молекул белков, определяя конформацию их гидрофобных участков;
Г) является метаболитом и непосредственным компонентом биохимических процессов, таких как фотосинтез (является донором электронов), дыхание (участвует в окислительных процессах цикла Кребса), гидролиз и синтез;
Д) высокая электронная и протонная проводимость определяют важную роль в мембранных процессах;
Е) основной компонент транспортной системы растений: как межклеточной – симпласт и апопласт, так и между органами – сосуды ксилемы и ситовидные трубки флоэмы;
Ж) является терморегулирующим фактором, защищая ткани от резких колебаний температуры благодаря высокой теплоемкости и большой удельной теплоте парообразования;
З) обеспечивает гидроскелет клеток и органов благодаря явлениям осмоса и тургора, от которых зависит характерная форма растительных тканей и органов.
Структура и свойства воды
Как физическое вещество, вода по многим показателям является аномальным.
Во-первых, плотность. Все вещества увеличивают обьем при нагревании, уменьшая при этом плотность. Однако при давлении 1 атм у воды в интервале от 0 до 4оС при увеличении температуры обьем уменьшается и максимальная плотность наблюдается при 4оС. При замерзании обьем воды резко возрастает на 11%, а при таянии льда при 0оС так же резко уменьшается. С увеличением давления температура замерзания воды понижается через каждые 130 атм на 1оС. Поэтому на больших глубинах при минусовых температурах вода в океане не замерзает.
Во-вторых, точки кипения и замерзания. При давлении 1 атм эти точки для воды составляют соответственно 0 и 100оС, что резко отличает воду от соединений ее группы.
В-третьих, теплоемкость. Величина теплоемкости, то есть количество теплоты, необходимое для повышения температуры на 1оС, в 5-30 раз выше, чем у других веществ. Кроме того, лишь у воды удельная теплоемкость с повышением температуры от 0 до 35оС падает, а затем начинает возрастать.
Поскольку теплоемкость песка в 5 раз меньше, чем у жидкой воды, то при одинаковом нагреве солнцем вода в водоеме нагревается в 5 раз слабее, чем песок на берегу, но во столько же раз дольше сохраняет теплоту. Высокая теплоемкость воды защищает растений от резкого повышения температуры при высокой температуре воздуха, а высокая теплота парообразования (2,3 кДж на 1 г) участвует в терморегуляции растений.
Поверхностное натяжение и прилипание. На поверхности воды из-за когезии ее молекул (силы сцепления между молекулами воды) создается поверхностное натяжение (0,72 мН/см), выше которого таковое только у ртути. Свойство же адгезии (прилипания) обнаруживается при подъеме воды против сил гравитации по сосудам.
В жидком состоянии каждая молекула воды является диполем (угол между Н составляет 105о) с тетраэдрическим распределением электронов вокруг атома кислорода и может взаимодействовать с четырьмя другими молекулами воды за счет электростатического взаимодействия атомов Н и О соседних молекул, то есть за счет водородной связи (энергия разрыва такой связи 18,84кДж/моль в отличие от ковалентной связи Н-О 460,4 кДж/моль).
Полярность молекулы воды обуславливает ее свойства растворителя. Хорошо растворяются в воде органические вещества с карбоксильными, карбонильными и с другими группами, с которыми вода образует водородные связи.
В растворах электролитов, содержащих ионы, структура воды существенно меняется. В электрическом поле катиона все ближайшие молекулы воды ориентируются отрицательными полюсами внутрь, а вокруг аниона внутрь направлены положительные полюсы молекул воды. Этот внутренний, прочно связанный с ионом слой молекул воды называют первичной гидратацией (сольватацией). Следующие слои воды также образуют относительно упорядоченную структуру, называемую вторичной гидратацией. Воду, связанную с ионами, называют также осмотически связанной. По мере повышения концентрации раствора (до 1,5 0 2 моль/л) вторичные гидратные оболочки ионов перекрываются и наблюдается переход к структуре кристаллогидрата.
В белках гидратация обусловлена взаимодействием молекул воды с гидрофильными (ионными и электронейтральными) и гидрофобными (неполярными) группами и ее иммобилизацией в замкнутых пространствах внутри макромолекул.
Формы воды в клетке.
В клетках и тканях различают две формы воды – свободную и связанную. Связанная вода подразделяется на А) связанную осмотически; б) коллоидно связанную, которая включает интрамицеллярную воду, находящуюся внутри коллоидной системы (в том числе и иммобилизационую воду) и интермицеллярную воду (находится на поверхности коллоидов и между ними); в) капиллярно связанную (находится в клеточных стенках и сосудах проводящей системы).
Вода удерживается в клетках за счет осмоса и набухания биоколлоидов. Клеточные стенки обладают значительной гигроскопичностью и удерживают воду в основном в силу высокой гидрофильности их пектиновых и целлюлозных компонентов. Они содержат две фракции воды: подвижную и малоподвижную. Вода, находящаяся в микрокапиллярных пространствах и та, которая удерживается водородными связями на микрофибриллах целлюлозы, считается малоподвижной, тогда как значительные количества воды в крупных капиллярных пространствах между микрофибриллами целлюлозы легко перемещаются. Содержание влаги в клеточных стенках тургресцентных клеток превышает 50%, а всего на долю клеточных стенок может приходится до ¼ всей внутриклеточной воды.
По сравнению с клеточными стенками цитоплазма содержит больше воды: ее содержание может достигать 95% от массы цитоплазмы, но это составляет лишь 1/20 всей клеточной воды. Основной вид гидрофильных коллоидов в цитоплазме – белки. Внутри белковых структур заключено значительное количество воды. Кроме того, цитоплазма содержит также сахара, соли, липиды и другие соединения, которые оказывают влияние на состояние содержащейся в ней воды.
Первое место по концентрации воды в клетке занимает вакуоль (98%, что составляет 3/4 всей воды). Вакуолярный сок содержит сахара, органические кислоты и их соли, неорганические катионы и анионы, ферменты, белки и другие азотистые соединения, слизи, танины, пигменты.
Осмотическое поглощение воды
Вода может поступать в клетки растений благодаря набуханию биоколлоидов, увеличению степени их гидратации. (например, сухие семена, помещенные в воду). Однако главным способом поступления воды в живые клетки является ее осмотическое поглощение.
Осмосом называется прохождение растворителя в раствор, отделенный от него полупроницаемой мембраной. Для разбавленных растворов осмотическое давление определяется концентрацией частиц (молекул, ионов) растворенного вещества. Согласно уравнению Вант-Гоффа:
P= i* c * RT, где
i- изотонический коэффициент, равный 1+ α(n-1), с – молярная концентрация раствора, R – универсальная газовая постоянная, Е – температура в Кельвинах.
При рассмотрении растительной клетки как осмотической системы обычно используют упрощенную модель, в которой полупроницаемой мембраной считается система, состоящая из плазмалеммы и тонопласта одновременно. Поскольку мембрана избирательно проницаема и вода проходит через нее значительно быстрее, чем вещества, растворенные в клеточном соке и цитоплазме, при помещении клетки в воду последняя по законам осмоса будет поступать внутрь клетки. Осмотическая концентрация вакуолярного сока для клеток корней составляет 0,3 – 1,2 МПа, а для клеток надземных органов – 1,0 – 2,6 Мпа. В клетках галофитов достигает 15 МПа.
Силу, с которой вода входит в клетку, называют сосущей силой S. Ее величина определяется осмотическим давлением клеточного сока Р и тургорным давлением (гидростатическим) Т, которое равно противодавлению клеточной стенки, возникающему при ее эластическом растяжении:
S= Р – Т
Когда клетка полностью насыщена водой (полностью тургресцентная) ее сосущая сила равна нулю, а тургорное давление равно осмотическому: S= 0; Р=Т. Состояние полного тургора наблюдается в клетках при достаточной влажности почвы и воздуха. Если подача воды к клетке уменьшается *(при усилении ветра, при недостатке влаги в почве), то вначале возникает водный дефицит в клеточных стенках, водный потенциал которых становится ниже, чем в вакуолях, и воды начинает перемещаться в клеточные стенки. Отток воды из вакуоли снижает тургорное давление в клетках и, следовательно, увеличивает их сосущую силу. При длительном недостатке влаги большинство клеток теряет тургор и растение подвядает. В этих условиях Т=0, S=Р.
Явление потери тургора клетками можно наблюдать экспериментально, помещая кусочки тканей в гипертонический раствор. В этом случае отток воды из клеток приводит к уменьшению объема протопластов ми их отделению от клеточных стенок (происходит плазмолиз). Пространство между стенкой и сократившимся протопластом заполняет наружный раствор. В условиях водного дефицита в молодых тканях резкое усиление потери воды, например при суховее, моет приводить к тому, что тургорное давление в клетках становится отрицательной величиной и протопласты, сокращаясь в объеме, не отделяются от клеточных стенок, а тянут их вслед за собой. Клетки и ткани сжимаются. Это явление называется циторриз.
Передвижение воды по растению
Наземный образ жизни растений обусловил необходимость создания внутри своего тела непрерывного восходящего тока воды. Этот ток начинается поглощением воды корнями, затем – передвижение ее по сосудам и, наконец, испарение воды листьями в результате транспирации.
Содержание доступной воды в почве определяется таким показателем как влажность устойчивого завядания, а максимальное содержание – полевой влагоемкостью. В среднем легкодоступная для растений влага удерживается в почве силой до 0.5 МПа, среднедоступная – до 1,0 -1,2 Мпа, а труднодоступная – до 2,5 – 3,0 Мпа.
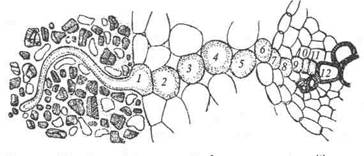
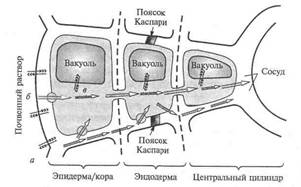 Зона наиболее интенсивного поглощения воды корневой системой – это зона корневых волосков, то есть зона всасывания (площадь которой в 22 раза больше испаряющей поверхности листьев). Вода поступает в цитоплазму клеток ризодермы по законам осмоса. Поскольку сопротивление клеточных стенок для воды значительно ниже, чем у цитоплазмы, более быстрый радиальный транспорт воды осуществляется через корень по апопласту. Однако на уровне эндодермы этот тип транспорта становится невозможным из-за непроницаемых для воды поясков Каспари. Поэтому происходит переключение с быстрого апопластного пути на медленный симпластный. Это своеобразный регулярный механизм подачи воды, так как диаметр стели, куда должна подаваться вода через эндодерму в 5-6 раз меньше диаметра поверхности корня.
Зона наиболее интенсивного поглощения воды корневой системой – это зона корневых волосков, то есть зона всасывания (площадь которой в 22 раза больше испаряющей поверхности листьев). Вода поступает в цитоплазму клеток ризодермы по законам осмоса. Поскольку сопротивление клеточных стенок для воды значительно ниже, чем у цитоплазмы, более быстрый радиальный транспорт воды осуществляется через корень по апопласту. Однако на уровне эндодермы этот тип транспорта становится невозможным из-за непроницаемых для воды поясков Каспари. Поэтому происходит переключение с быстрого апопластного пути на медленный симпластный. Это своеобразный регулярный механизм подачи воды, так как диаметр стели, куда должна подаваться вода через эндодерму в 5-6 раз меньше диаметра поверхности корня.
Рис.1 Путь води от корневого волоска 1 к сосудам коня 12через клетки паренхимы коры 2—6, эндодерму 7, перицикл 8 и паренхиму центрального цилиндра 9—11 и пути ближнего транспорта води: а — апопластный б — симпластный; в — трансвакуолярный
В сосуды ксилемы вода поступает, как и на первых этапах поглощения, благодаря осмотическому механизму. Осмотически активными веществами в сосудах и их клеточных стенках служат минеральные вещества и метаболиты, выделяемые активными ионными насосами, функционирующими в плазмалемме паренхимных клеток, окружающих сосуды. Высокая сосущая сила сосудов также определяется отсутствием противодавления со стороны лигнифицированных клеточных стенок.
Таким образом, в результате активной работы ионных насосов в корне и осмотическому (пассивному) поступлению воды в сосуды ксилемы в сосудах развивается гидростатическое давление, получившее название корневого давления. А механизм поднятия воды по растению вследствие развивающегося корневого давления называют нижним концевым двигателем.
Примером работы нижнего концевого двигателя служит «плач» растений – выделение ксилемного сока или пасоки из стебля весной у деревьев с еще нераспустившимися почками или явление гуттации – выделение на концах и зубчиках листьев капельно-жидкой воды при высокой влажности воздуха.
Движущей силой восходящего тока воды в проводящих элементах ксилемы является градиент сосущей силы клеток листьев и корня. В ответ на присасывающее действие транспирации вода поднимается вверх против сил гравитации благодаря силам когезии и адгезии со скоростью 20 см3 * ч-1 на 1 см2.
Транспирация – это физиологический процесс испарения воды растением. Основным органом транспирации является лист. Испарение воды с поверхности листьев или через специальные образования – устьица приводит к увеличению сосуще силы клеток листа. Что в свою очередь приводит к усилению поглощения клетками листа воды из ксилемы жилок и передвижению воды по ксилеме из корней в листья. Так работает верхний концевой двигатель. Причем он может работать и при полном отключении нижнего концевого двигателя, так как для его работы используется не метаболическая энергия, а энергия внешней среды – температура и движение воздуха.
Транспирация слагается из двух процессов: а) передвижения воды из листовых жилок в поверхностные слои стенок клеток мезофилла; б) испарения воды из клеточных стенок в межклеточные пространства и подустьичные полости с последующей диффузией в окружающую атмосферу через устьица (устьичная транспирация) или испарения из клеточных стенок эпидермиса путем кутикулярной транспирации (до 1/10 до ½ в зависимости от возраста).
Количество устьиц колеблется от 50 до 500 на 1 мм2 и хотя их площадь составляет всего 0,5 – 2,0% от общей поверхности листа испарение происходит почти со скоростью эвапорации (испарения со свободной поверхности).
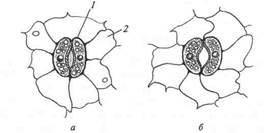
Рис.2. Устьичный аппарат:
1— замикающая клетка; 2 – клетка эпидермиса (а, б — разная степень открытости уститьц)
Открывание устьиц регулируется несколькими механизмами. Движущей силой, вызывающей открывание устьиц, является изменение тургора замыкающих клеток. По мере того, как замыкающая клетка осмотически поглощает воду, более тонкая и эластичная часть ее клеточной стенки, удаленная от щели, растягивается. Поскольку более толстый и менее эластичный участок стенки, окаймляющий щель, растягивается слабее, замыкающие клетки принимают полукруглую форму, в результате чего устьица раскрываются. Если движения зависят от изменения в самих замыкающих клетках, то они называются гидроактивными (фотоактивное открытие- на накопление сахаров в результате фотосинтеза или гидроактивное открытие в результате повышения концентрации К+ в замыкающих клетках, гидроактивное закрывание в ответ на снижения тургора в результате превышения транспирации над поглощением, то есть по мере развития водного дефицита и повышения концентрации АБК). Если же движение определяется изменениями в клетках, окружающих устьичные, то они называются гидропассивнми (гидропассивное сдавливание).
Интенсивность транспирации – количество испаренной воды за 1 ч на единицу площади или на 1 г сухого вещества (обычно 15-250 г*м2*ч-1 днем и 1-20 ночью).
Продуктивность транспирации _ количество граммов сухого вещества, образуемых при расходовании 1000 г воды (в умеренном климате 1-8 г).
Транспирационный коэффициент – число граммов воды, израсходованной при накоплении 1 г сухих веществ (от 125 до 1000). Следовательно, на синтез веществ своего тела растение использует лишь 0,2% пропускаемой воды, остальные 99,8% тратятся на испарение.
Таким образом, постоянный ток воды от корневой системы к надземным частям: 1) транспортирует минеральные вещества; 2) обеспечивает нормальное водоснабжение и поддержание тургора; 3) способ защиты от перегрева.
Экологические группы растений по отношению к воде (гигро-, ксеро- и мезофиты) – самостоятельно.

