




Эта группа жирных кислот окисляется по такому же механизму, что и обычные жирные кислоты с четным числом атомов углерода. После поступления в клетку они активируются с образованием ацил-КоА и потреблением АТФ, затем транспортируются в митохондрии с помощью карнитинового челнока, где разрушаются в результате β-окисления.
Остающийся пропионил-КоА карбоксилируется пропионил-КоА-карбоксилазой с образованием метилмалонил-КоА, который после изомеризации превращается в сукцинил-СоА.
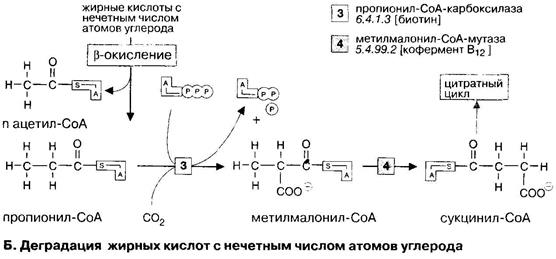
В этих реакциях принимают участие различные коферменты: карбоксилирование происходит с участием биотина, а изомеризация - с участием кофермента витамина В12(5’дезоксиаденозилкобаламина).
Сукцинил-КоА является промежуточным метаболитом цитратного цикла и после превращения в оксалоацетат включается в глюконеогенез. Таким образом,из конечного продукта деградации жирных кислот с нечетным числом атомов углерода - пропионил-СоА - синтезируется глюкоза. Напротив, образующийся при β- окислении жирных кислот ацетил-КоА не может использоваться для глюконеогенеза, так как оба углеродных атома ацетильного остатка на пути к оксалоацетату превращаются в СО2.
Итак, для полной деградации длинноцепочечной жирной кислоты цикл должен многократно повторяться; например, для С17Н35-S-КоА (С18) необходимы восемь циклов. Образующийся ацетил-КоА далее может метаболизироваться двумя путями:
1. Взаимодействовать с оксалоацетатом с образованием цитрата, промежуточного метаболита цитратного цикла; при этом ацетильный остаток полностью окисляется в цикле Кребса до СО2.
2. При избытке ацетил-КоА в гепатоцитах образуются кетоновые тела.
Биосинтез кетоновых тел
В том случае, если количество образовавшегося ацетил-КоА превосходит энергетическую потребность гепатоцитов, что наблюдается при высоком содержании жирных кислот в плазме крови (типичные случаи - голодание и сахарный диабет), то в гепатоцитах синтезируются так называемые кетоновые тела.
При высокой концентрации ацетил-КоА в митохондриях гепатоцитов происходит конденсация двух молекул ацетил-КоА с образованием ацетоацетил-КоА.

Присоединение еще одной ацетильной группы приводит к 3-гидрокси-3-метилглутарил-КоА (ГМГ-КоА).

ГМГ-КоА после отщепления ацетил-КоА превращается в ацетоуксусную кислоту (ацетоацетат) (цикл Линена).
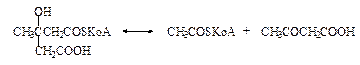
При восстановлении ацетоуксусной кислоты НАДН получается З-гидроксибутират, а при неферментативном декарбоксиливании ацетоуксусной кислоты - ацетон.

Эти три соединения (ацетон, ацетоуксусная кислота и 3-гидроксимасляная кислота) принято называть «кетоновыми телами», что не совсем правильно, поскольку в 3-гидроксимасляной кислоте отсутствует кетогруппа!
Кетоновые тела поступают из печени в кровь. Они хорошо растворимы в воде и следовательно в крови. Концентрация кетоновых тел в крови возрастает в период голодания. Наряду с жирными кислотами 3-гидроксибутират и ацетоацетат в этот период являются основными энергоносителями. Ацетон, не имеющий метаболической ценности, удаляется через легкие. После 1-2 недель голодания кетоновые тела начинают использоваться в качестве источника энергии нервными тканями. Однако при этом для обеспечения работы цитратного цикла все же необходимо минимальное количество глюкозы.
Если биосинтез кетоновых тел превышает потребности организма, они накапливаются в крови (кетонемия) и, наконец, выводятся с мочой (кетонурия). Оба феномена наблюдаются вовремя длительного голодания (углеводная недостаточность) и при заболевании диабетом.Хотя 3-гидроксимасляная кислота является слабой кислотой (рКа примерно 4), возрастание концентрации кетоновых тел вызывает изменение рН в крови (кетоацидоз). Кетонурия и кетоацидоз могут привести к нарушению ионного гомеостаза и как следствие потере сознания (кетоацидозной коме) и, следовательно, опасны для жизни. Общая схема образования кетоновых тел приведена ниже.


