




СЛАЙД 1
Лекция №4. Гуморальные факторы врожденного иммунитета
Содержание
1. Система комплемента
2. Белки острой фазы воспаления
3. Биогенные амимны
4. Липидные медиаторы
5. Цитокины
6. Интерфероны
СЛАЙД 2
Гуморальная составляющая врожденного иммунитета представлена несколькими взаимосвязанными системами — системой комплемента, цитокиновой сетью, бактерицидными пептидами, а также гуморальными системами, связанными с воспалением.
Действие большинства этих систем подчиняется одному из двух принципов — каскада и сети. По каскадному принципу функционирует система комплемента, при активации которой происходит последовательное вовлечение факторов. При этом эффекты каскадных реакций проявляются не только в конце активационного пути, но и на промежуточных стадиях.
Принцип сети характерен для системы цитокинов и предполагает возможность одновременного функционирования различных компонентов системы. Основа функционирования такой системы — тесная взаимосвязь, взаимное влияние и значительная степень взаимозаменяемости компонентов сети.
СЛАЙД 3
Комплемент – сложный белковый комплекс сыворотки крови.
Система комплемента состоит из 30 белков (компонентов, или фракций, системы комплемента).
Активируется система комплемента за счет каскадного процесса: продукт предыдущей реакции исполняет роль катализатора последующей реакции. Причем при активации фракции компонента происходит, у первых пяти компонентов, ее расщепление. Продукты этого расщепления и обозначаются как активные фракции системы комплемента.
1. Больший из фрагментов (обозначаемый буквой b), образовавшихся при расщеплении неактивной фракции, остается на поверхности клетки – активация комплемента всегда происходит на поверхности микробной клетки, но не собственных эукариотических клеток. Этот фрагмент приобретает свойства фермента и способность воздействовать на последующий компонент, активируя его
2. Меньший фрагмент (обозначается буквой a) является растворимым и «уходит» в жидкую фазу, т.е. в сыворотку крови.
Фракции системы комплемента обозначаются по-разному.
1. Девять – открытых первыми – белков системы комплемента обозначаются буквой С (от английского слова complement) с соответствующей цифрой.
2. Остальные фракции системы комплемента обозначаются другими латинскими буквами или их сочетаниями.
СЛАЙД 4
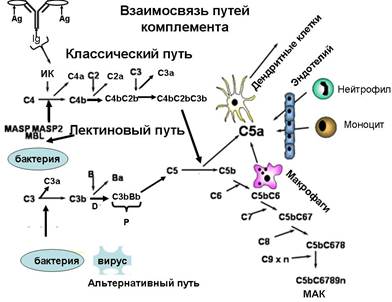
Пути активации комплемента
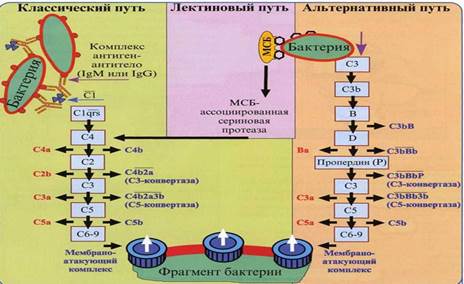
Существуют три пути активации комплемента: классический, лектиновый и альтернативный.
СЛАЙД 5
1. Классический путь активации комплемента является основным. Участие в этом пути активации комплемента – главная функция антител.
Активацию комплемента по классическому пути запускает иммунный комплекс: комплекс антигена с иммуноглобулином (класса G или М). Место антитела может «занять» С-реактивный белок – такой комплекс также активирует комплемент по классическому пути.
Классический путь активации комплемента осуществляется следующим образом.
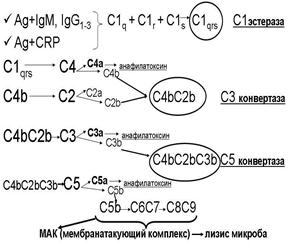
а. Сначала активируется фракция С1: она собирается из трех субфракций (C1q, C1r, C1s) и превращается в фермент С1-эстеразу (С1qrs).
б. С1-эстераза расщепляет фракцию С4.
в. Активная фракция С4b ковалентно связывается с поверхностью микробных клеток - здесь присоединяет к себе фракцию С2.
г. Фракция С2 в комплексе с фракцией С4b расщепляется С1-эстеразой с образованием активной фракции С2b.
д. Активные фракции С4b и С2b в один комплекс – С4bС2b – обладающий ферментативной активностью. Это так называемая С3-конвертаза классического пути.
е. С3-конвертаза расщепляет фракцию С3, нарабатываю большие количества активной фракции С3b.
ж. Активная фракция С3b присоединяется к комплексу С4bС2b и превращает его в С5-конвертазу (С4bС2bС3b).
з. С5-конвертаза расщепляет фракцию С5.
и. Появившаяся в результате этого активная фракция С5b присоединяет фракцию С6.
к. Комплекс С5bС6 присоединяет фракцию С7.
л. Комплекс С5bС6С7 встраивается в фосфолипидный бислой мембраны микробной клетки.
м. К этому комплексу присоединяется белок С8 и белок С9. Данный полимер формирует в мембране микробной клетки пору диаметром около 10 нм, что приводит к лизису микроба (так как на его поверхности образуется множество таких пор – «деятельность» одной единицы С3-конвертазы приводит к появлению около 1000 пор). Комплекс С5bС6С7С8С9, образующийся в результате активации комплемента, называется мемранатакующим комплексом (МАК).
СЛАЙД 6
2. Лектиновый путь активации комплемента запускается комплексом нормального белка сыворотки крови – маннансвязывающего лектина (МСЛ) – с углеводами поверхностных структур микробных клеток (с остатками маннозы).
СЛАЙД 7
3. Альтернативный путь активации комплемента начинается с ковалентного связывания активной фракции С3b – которая всегда присутствует в сыворотке крови в результате постоянно протекающего здесь спонтанного расщепления фракции С3 – с поверхностными молекулами не всех, но некоторых микроорганизмов.
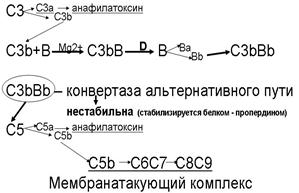
1. Дальнейшие события развиваются следующим образом.
а. С3b связывает фактор В, образуя комплекс С3bВ.
б. В связанном с С3b виде фактор В выступает в качестве субстрата для фактора D (сывороточной сериновой протеазы), которая расщепляет его с образованием активного комплекса С3bВb. Этот комплекс обладает ферментативной активностью, структурно и функционально гомологичен С3-конвертазе классического пути (С4bС2b) и называется С3-конвертазой альтернативного пути.
в. Сама по себе С3-конвертаза альтернативного пути нестабильна. Чтобы альтернативный путь активации комплемента успешно продолжался этот фермент стабилизируется фактором Р (пропердином).
г. То, что происходит дальше, аналогично классическому пути активации комплемента.
2. Основное функциональное отличие альтернативного пути активации комплемента, по сравнению с классическим, заключается в быстроте ответа на патоген: так как не требуется время для накопления специфических антител и образования иммунных комплексов.
Важно понимать, что и классический и альтернативный пути активации комплемента действуют параллельно, еще и амплифицируя (т.е. усиливая) друг друга. Другими словами комплемент активируется не «или по классическому или по альтернативному», а «и по классическому и по альтернативному» путям активации. Это, еще и с добавлением лектинового пути активации, – единый процесс, разные составляющие которого могут просто проявляться в разной степени.
СЛАЙД 8
Функции системы комплемента
Система комплемента играет очень важную роль в защите макроорганизма от патогенов.
1. Система комплемента участвует в инактивации микроорганизмов, в т.ч. опосредует действие на микробы антител.
2. Активные фракции системы комплемента активируют фагоцитоз (опсонины - С3b и C5b).
3. Активные фракции системы комплемента принимают участие в формировании воспалительной реакции.
СЛАЙД 9
Активные фракции комплемента С3а и С5а называются анафилотоксинами, так как участвуют, помимо прочего, в аллергической реакции, называемой анафилаксия. Наиболее сильным анафилотоксином является С5а. Анафилотоксины действуют на разные клетки и ткани макроорганизма.
1. Действие их на тучные клетки вызывает дегрануляцию последних.
2. Анафилотоксины действуют также на гладкие мышцы, вызывая их сокращение.
3. Действуют они и на стенку сосуда: вызывают активацию эндотелия и повышение его проницаемости, что создает условия для экстравазации (выхода) из сосудистого русла жидкости и клеток крови в ходе развития воспалительной реакции.
Корме того, анафилотоксины являются иммуномодуляторами, т.е. они выступают в роли регуляторов иммунного ответа.
1. С3а выступает в роли иммуносупрессора (т.е. подавляет иммунный ответ).
2. С5а является иммуностимулятором (т.е. усиливает иммунный ответ).
СЛАЙД 10
Белки острой фазы
Некоторые гуморальные реакции врожденного иммунитета по своему назначению аналогичны реакциям адаптивного иммунитета и могут рассматриваться как их эволюционные предшественники. Такие реакции врожденного иммунитета имеют преимущество перед адаптивным иммунитетом в быстроте развития, однако недостаток их заключается в отсутствии специфичности в отношении антигенов. Пару сходных по результатам реакций врожденного и адаптивного иммунитета мы рассмотрели выше в разделе, посвященном комплементу (альтернативная и классическая активация комплемента). Другой пример будет рассмотрен в данном разделе: белки острой фазы в ускоренном и упрощенном варианте воспроизводят некоторые эффекты антител.
Белки (реактанты) острой фазы представляют группу протеинов, секретируемых гепатоцитами. При воспалении продукция белков острой фазы изменяется. При усилении синтеза белки называют положительными, а при понижении синтеза — отрицательными реактантнами острой фазы воспаления.
Динамика и выраженность изменений сывороточной концентрации различных белков острой фазы при развитии воспаления неодинакова: концентарция С-реактивного белка и сывороточного амилоида Р возрастает очень сильно (в десятки тысяч раз) — быстро и кратковременно (практически нормализуется к концу 1-й недели); уровни гаптоглобина и фибриногена возрастают слабее (в сотни раз) соответственно на 2-й и 3-й неделях воспалительной реакции. В данной презентации будут рассмотрены только положительные реактанты, участвующие в иммунных процессах.
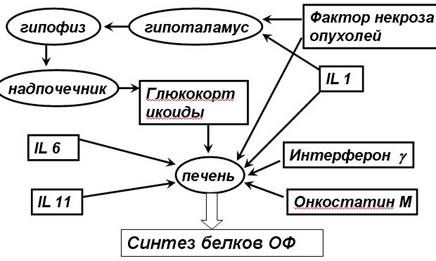
СЛАЙД 11
Согласно выполняемым функциям выделяют несколько групп белков острой фазы.
К транспортным белкам относят преальбумин, альбумин, орозомукоид, липокалины, гаптоглобин, трансферрин, маннозасвязывающий и ретинолсвязывающий белки и т.д. Они играют роль переносчиков метаболитов, ионов металлов, физиологически активных факторов. Роль факторов этой группы существенно возрастает и качественно изменяется при воспалении.
Другую группу образуют протеазы (трипсиноген, эластаза, катепсины, гранзимы, триптазы, химазы, металлопротеиназы), активация которых необходима для формирования многих медиаторов воспаления, а также для осуществления эффекторных функций, в частности киллерной. Активация протеаз (трипсина, химотрипсина, эластазы, металлопротеиназ) уравновешивается накоплением их ингибиторов. α2-Макроглобулин участвует в подавлении активности протеаз разных групп.
Помимо перечисленных, к белкам острой фазы относят факторы коагуляции и фибринолиза, а также белки межклеточного матрикса (например, коллагены, эластины, фибронектин) и даже белки системы комплемента.
СЛАЙД 12
Пентраксины. Наиболее полно проявляют свойства реактантов острой фазы белки семейства пентраксинов: в первые 2-3 сут развития воспаления их концентрация в крови повышается на 4 порядка.
С-реактивный белок и сывороточный амилоид Р образуются и секретируются гепатоцитами. Основной индуктор их синтеза — IL-6. Белок PTX3 вырабатывают миелоидные (макрофаги, дендритные клетки), эпителиальные клетки и фибробласты в ответ на стимуляцию через TLR, а также под действием провоспалительных цитокинов (например, IL-1β, TNFα).
Концентрация пентраксинов в сыворотке резко возрастает при воспалении: С-реактивного белка и сывороточного амилоида Р — с 1 мкг/мл до 1–2 мг/мл (т.е. в 1000 раз), РТХ3 — с 25 до 200–800 нг/мл. Пик концентрации достигается через 6–8 ч после индукции воспаления. Для пентраксинов характерна способность связываться с самыми разнообразными молекулами.
С-реактивный белок был впервые идентифицирован благодаря его способности связывать полисахарид С (Streptococcus рneumoniae), что и определило его название. Пентраксины взаимодействуют и с множеством других молекул: C1q, бактериальными полисахаридами, фосфорилхолином, гистонами, ДНК, полиэлектролитами, цитокинами, белками межклеточного матрикса, сывороточными липопротеинами, компонентами комплемента, друг с другом, а также с ионами Са 2+ и других металлов.
Для всех рассматриваемых пентраксинов существуют высокоаффинные рецепторы на миелоидных, лимфоидных, эпителиальных и других клетках. Кроме того, эта группа белков острой фазы обладает достаточно высоким сродством к таким рецепторам, как FcγRI и FcγRII. Многочисленность молекул, с которыми взаимодействуют пентраксины, определяет широкое разнообразие их функций.
Распознавание и связывание пентраксинами PAMP дает основание рассматривать их как вариант растворимых патогенраспознающих рецепторов.
К наиболее важным функциям пентраксинов относят их участие в реакциях врожденного иммунитета в качестве факторов, запускающих активацию комплемента через C1q и участвующих в опсонизации микроорганизмов.
Комплементактивирующая и опсонизирующая способность пентраксинов делает их своеобразными «протоантителами», частично выполняющими функции антител на начальном этапе иммунного ответа, когда истинные адаптивные антитела еще не успели выработаться.
Роль пентраксинов во врожденном иммунитете заключается также в активации нейтрофилов и моноцитов/макрофагов, регуляции синтеза цитокинов и проявлении хемотаксической активности по отношению к нейтрофилам. Помимо участия в реакциях врожденного иммунитета пентраксины регулируют функции межклеточного матрикса при воспалении, контроле апоптоза и элиминации апоптотических клеток.
СЛАЙД 13
Биогенные амины
К этой группе медиаторов относят гистамин и серотонин, содержащиеся в гранулах тучных клеток. Освобождаясь при дегрануляции, эти амины вызывают разнообразные эффекты, играющие ключевую роль в развитии ранних проявлений гиперчувствительности немедленного типа.
Гистамин (5-β-имидазолилэтиламин) — главный медиатор аллергии. Он образуется из гистидина под влиянием фермента гистидиндекарбоксилазы.
Поскольку гистамин содержится в гранулах тучных клеток в готовом виде, а процесс дегрануляции происходит быстро, гистамин очень рано появляется в очаге аллергического поражения, причем сразу в большой концентрации, что определяет проявления немедленной гиперчувствительности. Гистамин быстро метаболизируется (95% за 1 мин) с участием 2 ферментов — гистамин-N-метилтрансферазы и диаминооксидазы (гистаминазы); при этом образуется (в соотношении примерно 2:1) соответственно N-метилгистамин и имидазолацетат.
Известно 4 разновидности рецепторов для гистамина Н1-Н4. При аллергических процессах гистамин действует преимущественно на гладкие мыщцы и эндотелий сосудов, связываясь с их Н1-рецепторами. Эти рецепторы поставляют активационный сигнал, опосредованный превращениями фосфоинозитидов с образованием диацилглицерола и мобилизацией Са2+.
Указанные эффекты частично обусловлены образованием в клетках (мишенях гистамина) оксида азота и простациклина. Действуя на нервные окончания, гистамин вызывает ощущение зуда, характерного для аллергических проявлений в коже.
У человека гистамин играет важную роль в развитии кожной гиперемии и аллергического ринита. Менее очевидно его участие в развитии общих аллергических реакций и бронхиальной астмы. В то же время через Н2-рецепторы гистамин и родственные вещества оказывают регуляторное действие, иногда уменьшающее проявления воспаления, ослабляя хемотаксис нейтрофилов и выброс ими лизосомных ферментов, а также высвобождение самого гистамина.
Через Н2-рецепторы гистамин действует на сердце, секреторные клетки желудка, подавляет пролиферацию и цитотоксическую активность лимфоцитов, а также секрецию ими цитокинов. Большинство этих эффектов опосредовано активацией аденилатциклазы и повышением внутриклеточного уровня цАМФ.
Данные об относительной роли различных рецепторов гистамина в реализации его действия очень важны, поскольку многие антиаллергические препараты представляют собой блокаторы Н1 (но не Н2 и других) рецепторов гистамина.
СЛАЙД 14
Липидные медиаторы.
Важную роль в регуляции иммунных процессов, а также в развитии аллергических реакций играют гуморальные факторы липидной природы. Наиболее многочислены и важны из них эйкозаноиды.
Эйкозаноиды — продукты метаболизма арахидоновой кислоты — жирной полиненасыщенной кислоты, молекула которой содержит 20 атомов углерода и 4 ненасыщенные связи. Арахидоновая кислота образуется из мембранных фосфолипидов как прямой продукт действия фосфолипазы А (PLA) или косвенный продукт превращений, опосредованных PLC.
Образование арахидоновой кислоты или эйкозаноидов происходит при активации различных типов клеток, особенно участвующих в развитии воспаления, в частности аллергического: эндотелиальных и тучных клеток, базофилов, моноцитов и макрофагов.
Метаболизм арахидоновой кислоты может проходить по 2 путям — катализироваться циклооксигеназой или 5’-липоксигеназой. Циклооксигеназный путь приводит к образованию простагландинов и тромбоксанов из нестабильных промежуточных продуктов — эндоперекисных простагландинов G2 и H2, а липоксигеназный — к образованию лейкотриенов и 5-гидроксиэйкозатетраеноата через промежуточные продукты (5-гидроперокси-6,8,11,14-эйкозатетраеновую кислоту и лейкотриен А4), а также липоксинов — продуктов двойной липоксигенации (под действием двух липоксигеназ — см. далее).
Простагландины и лейкотриены во многих отношениях проявляют альтернативные физиологические эффекты, несмотря на то, что внутри этих групп существуют значительные различия в активности.
Общее свойство этих групп факторов — преобладающее действие на стенку сосудов и гладкие мышцы, а также хемотаксический эффект. Эти эффекты реализуются при взаимодействии эйкозаноидов со специфическими рецепторами на поверхности клеток. Некоторые представители семейства эйкозаноидов усиливают действие других вазоактивных и хемотаксических факторов, например, анафилатоксинов (С3а, С5а).
СЛАЙД 15
Лейкотриены (LT) — С20 -жирные кислоты, молекула которых в положении 5 содержит ОН-группу, а в положении 6 — боковые серосодержащие цепи, например глутатион.
Выделяют 2 группы лейкотриенов:
-одна из них включает лейкотриены С4, D4 и Е4, называемые цистеиниллей-котриенами (Cys-LT),
-во вторую входит один фактор — лейкотриен B4.
Лейкотриены образуются и секретируются в течение 5–10 мин после активации тучных клеток или базофилов.
Лейкотриен C4 присутствует в жидкой фазе в течение 3–5 мин, при этом он превращается в лейкотриен D4. Лейкотриен D4 существует в последующие 15 мин, медленно превращаясь в лейкотриен E4.
Лейкотриены оказывают свое действие через рецепторы, относящиеся к группе пуриновых рецепторов семейства родопсиноподобных рецепторов, 7-кратно пронизывающих мембрану и связанных с протеином G.
Рецепторы лейкотриенов экспрессируются на клетках селезен-ки, лейкоцитах крови, кроме того, CysLT-R1 представлен на макрофагах, клетках кишечника, воздухоносного эпителия, а CysLT-R2 — на клетках надпочечников и головного мозга.
Цистеиниловые лейкотриены (особенно лейкотриен D4) вызывают спазм гладкой мускулатуры и регулируют локальный кровоток, снижая артериальное давление. Цистеиниловые лейкотриены — медиаторы аллергических реакций, в частности, медленной фазы бронхоспазма при бронхиальной астме.
Кроме того, они подавляют пролиферацию лимфоцитов и способствуют их дифференцировке.
Ранее комплекс этих факторов (лейкотриены C4, D4 и E4) называли медленнореагирующей субстанцией А. Лейкотриен B4 (дигидроксиэйкозатетраеновая кислота) проявляет хемотаксическое и активирующее действие преимущественно в отношении моноцитов, макрофагов, нейтрофилов, эозинофилов и даже Т-клеток.
Еще один продукт липоксигеназного пути — 5-гидроксиэйкозатетраеноат — менее активен, чем лейкотриены, но может служить хемоаттрактантом и активатором нейтрофилов и тучных клеток.
СЛАЙД 16
Простагландины (PG) — С20 -жирные кислоты, молекула которых содержит циклопентановое кольцо.
Варианты простагландинов, отличающиеся по типу и положению замещающих групп (окси-, гидрокси-), обозначаются различными буквами; цифры в названии означают число ненасыщенных связей в молекуле.
Простагландины накапливаются в очаге воспаления позже кининов и гистамина, несколько позже лейкотриенов, но одновременно с монокинами (через 6–24 ч после запуска воспаления).
Помимо вазоактивного и хемотаксического эффекта, достигаемого в кооперации с другими факторами, простагландины (особенно простагландин E2) оказывают регулирующее действие при воспалительных и иммунных процессах.
Экзогенный простагландин E2 вызывает некоторые проявления воспалительной реакции, но подавляет иммунный ответ и аллергические реакции.
Так, простагландин E2 снижает цитотоксическую активность макрофагов, нейтрофилов и лимфоцитов, пролиферацию лимфоцитов, выработку этими клетками цитокинов.
Он способствует дифференцировке незрелых лимфоцитов и клеток других кроветворных рядов.
Некоторые эффекты простагландина Е2 связаны с повышением уровня внутриклеточного цАМФ.
Простагландины E2 и D2 подавляют агрегацию тромбоцитов; простагландины F2 и D2 вызывают сокращение гладкой мускулатуры бронхов, тогда как простагландин E2 расслабляет ее.
СЛАЙД 17
Тромбоксан А2 (ТХА2) — С20 -жирная кислота; в его молекуле есть 6-членное кислородсодержащее кольцо.
Это очень нестабильная молекула (время полужизни — 30 с), превращающаяся в неактивный тромбоксан В2.
Тромбоксан А2 вызывает сужение сосудов и бронхов, агрегацию тромбоцитов с высвобождением из них ферментов и других активных факторов, способствующих митогенезу лимфоцитов.
Другой продукт циклоксигеназного пути — простагландин I2 (простациклин) — тоже нестабилен. Он проявляет свое действие через цАМФ, сильно расширяет сосуды, увеличивает их проницаемость, ингибирует агрегацию тромбоцитов.
Наряду с пептидным фактором брадикинином простациклин вызывает ощущение боли при воспалении.
СЛАЙД 18
Цитокины

