




Центральные генераторы паттернов, управляющие каждым из этих видов активности, обладают определенными общими свойствами. В их основе лежит набор циклических скоординированных во времени сигналов, которые генерируются кластером взаимосвязанных нейронов. Эти основные сигналы используются для управления несколькими сотнями мышц, каждая из которых с точностью сокращается или расслабляется во время определенной фазы цикла. Ритмические паттерны генерируются в спинном мозге, а нейроны ствола головного мозга контролируют возникновение паттерна и его скорость. головного

Каким образом нейронные цепи генерируют ритмические паттерны активности? На этот вопрос нет однозначного ответа, поскольку различные нейронные цепи используют различные механизмы. Самые простые генераторы паттернов — командные нейроны, которые вследствие особенностей своих мембран обладают пейсмейкерными свойствами, аналогично свойствам мышечных клеток сердца и гладкомышечных клеток. Даже будучи изолированными экспериментальным путем от других нейронов, нейроны-пейсмейкеры способны генерировать ритмичные паттерны только на основе проводимости своих мембран. Легко представить, каким образом истинные нейроны-пейсмекеры могут функционировать в качестве первичных водителей ритма для мотонейронов, которые в свою очередь управляют цикличными поведенческими процессами. Однако, хотя нейроны-пейсмейкеры могут вносить вклад в работу некоторых центральных генераторов паттернов, в случае позвоночных животных, по-видимому, не только они ответственны за генерацию ритмов. Нейроны-пейсмейкеры находятся внутри взаимосвязанных нейронных цепей, и именно комбинация настоящих пейсмейкерных свойств нейронов и синаптических контактов между нейронами лежит в основе генерации ритмических паттернов.

A Hypothetical Pattern Generator consisting of two pacemaker cells (1
and 2) that excite flexor and extensor motoneuron pools, respectively. The
pacemaker cells discharge with a spontaneous rhythm and are coupled by
inhibitory interneurons (black) that ensure that their discharge is out of phase.
When pacemaker 1 fires, the flexor motoneuron is activated and pacemaker 2 is
suppressed. When pacemaker 1 falls silent pacemaker 2 can fire, maintaining the
silence of pacemaker 1 and activating the extensor motoneuron. The upper two
traces represent intracellular records from pacemakers 1 and 2; the lower traces
represent extracellular recordings from flexor (F) and extensor (E) motoneurons.
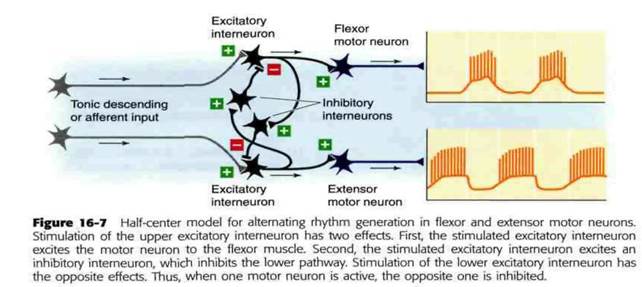
Нейронные цепи, в состав которых не входят нейроны-пейсмекеры (командные нейроны), также могут генерировать ритмические выходные сигналы. В 1911 г. Грэм Браун (Graham Brown) предложил модель нейронной цепи, генерирующей двигательные паттерны для локомоции. В основе полуцентрической модели Брауна лежит набор возбуждающих и тормозных интернейронов, расположенных таким образом, чтобы реципрокным путём ингибировать друг друга. Полуцентры представляют собой две половины нейронной цепи, каждая из которых командует одной парой мышц-антагонистов. Чтобы такая нейронная цепь работала, возбуждающие интернейроны нуждаются в тоническом стимуле. Такой стимул может поступить от аксонов, которые берут начало за пределами нейронной цепи, или возникнуть вследствие собственной возбудимости этих нейронов. Кроме того, некий встроенный механизм должен ограничивать продолжительность тормозной активности интернейронов, чтобы возбудимость циклически передавалось от одного полуцентра к другому. Следует отметить, что обратная связь от рецепторов мышц не нужна для того, чтобы ритмы продолжались неограниченно долго. Исследования, проведенные на более чем 50 двигательных нейронных цепях позвоночных и беспозвоночных животных, подтвердили, что генерация ритмов может длиться в отсутствие сенсорной информации.
Особенности центральных программ:
· Сенсорная обратная связь для изменения двигательных ритмов (одинаково важна обратная связь от рецепторов мышц, суставов и кожи).
· Координация активности генераторов паттернов в различных сегментах спинного мозга. Например: каждый сегмент спинного мозга должен дать команду к сокращению «своим» мышцам немного позднее, чем сегмент, расположенный перед ним, причем запаздывание составляет приблизительно 1% полного цикла сокращения мышц во время нормального плавания по направлению вперед. При некоторых обстоятельствах минога использует обратную последовательность координации действий между сегментами спинного мозга, что позволяет ей тогда плыть задом наперед.
· Поддержание реципрокной связи между спинальными генераторами и управляющими центрами ствола мозга. При этом не только ствол мозга использует многочисленные пути и медиаторы для модуляции работы генераторов паттернов в спинном мозге, но и генераторы паттернов спинного мозга информируют ствол мозга о своей деятельности.

