




Однако какими бы определенными ни были эти результаты, они оставляют нерешенным первоначальный вопрос — вопрос о том, как возникает голод. Эта проблема оставалась нерешенной до тех пор, пока не были проведены некоторые эксперименты. О;;;1 внесли известную ясность в этот вопрос.
Я занимался изучением функций не ствола мозга, а височной доли полушарий и обнаружил, что расстройства в акте еды (рис. Х-2), возникающие при удалении значительной части этой доли, могут быть вызваны также удалением небольшой части медиальной поверхности ее полюса — амигдалы, структуры лим-бической системы, относимой обычно к одному из базальных ганглиев (рис. Х-3). Мои опыты были проведены на обезьянах, пищевое поведение которых особенно трудно для изучения. Обезьяны сохраняют пищу в защечных мешках; они очищают и растирают предложенную им пищу, пробуют и выбрасывают ее, когда насытятся. И все же предварительные количественные результаты показали, что удаление амигдалы удваивает количество поглощенной пищи.
Мои коллеги по отделению физиологии в Йельском университете, которые научились вызывать ожирение у крыс, разрушая гипоталамический механизм «насыщения», и особенно Дж. Бро-бек заинтересовались этой проблемой и вместе с Б. Анандом, ординатором из Индии, начали над нею работать. Мы спланиро-
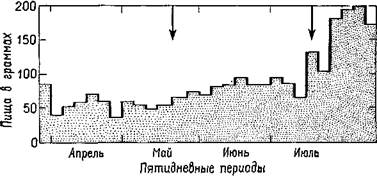
Рис Х-2 Влияние двустороннего удаления амигдалы на поглощение пищи обезьяной, содержащейся на свободной диете Стрелки обозначают операцию резекцию первой, а затем второй амигдалы (Pribram and Bagshow, 1953)
вали свои действия: я должен был производить хирургические повреждения, тогда как упомянутые сотрудники вводили в амигдалу зонд для разрушения ее электрическим током Мы создавали такие условия, которые, как мы полагали, должны были дать ответ на вопрос, связано ли расстройство питания с изменением вкусовых ощущений (последнее измерялось путем относительного поглощения горького и сладкого растворов и смеси из опилок и колесной мази и жирного пюре, по виду и консистенции похожего на эту смесь, но вполне съедобного для животных) или с некоторым нарушением обмена веществ (у животных регулярно измерялась температура и вес, а также двигательная активность животного, помещенного в специальное колесо).
Нашим руководителем, благословившим эти исследования, должен был стать Р. Берне. Все наши тщательные и кропотливые измерения оказались напрасными: результаты опытов были драматичны и видны невооруженным глазом Прежде всего половина крыс погибла не от операции, а до нее, потому что они пили очень много горького раствора и ели так много тавота и опилок, что серьезно заболели, получив хинное отравление. Дикие крысы, как известно, различают, что им полезно (и те из крыс, которые оставались в живых, также различали это, потому что они никогда не прикасались снова к вредным веществам), но специально выведенные в лаборатории белые крысы не обладали такой способностью И хотя я действительно обнаружил, что после удаления амигдалы крысы снова начинают пить горький раствор и поглощать опилки с тавотом, у меня не было ощущения, что эксперимент завершен А поскольку этот опыт также не дал ответа на вопросы, относящиеся к изучаемой проблеме, мы никогда не сообщали о его результатах.
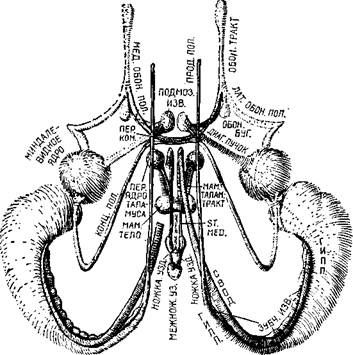
Рис Х-3 Реконструкция системы связей между лимбическими
(амигдала и гиппокамп) и стволовыми (гипоталамус) структурами
мозга (Krieg, 1966).
С моими коллегами, Анандом и Бробеком, случилось даже худшее или, возможно, лучшее в зависимости от того, как на это посмотреть Почти все их крысы погибали после стереотаксической операции — и снова не от самого хирургического вмешательства, а от того, что животные прекращали одновременно есть и пить. Эти результаты были противоположны тем, которые я получил на обезьянах, хотя временный послеоперационный отказ от пищи и питья часто имел место и у них. На этой стадии почти год спустя после начала экспериментов мы, в общем, не мо1Л1г считать полученные результаты удовлетворительными. Поэтому Ананд (который вскоре должен был вернуться в Индию) взял на себя тяжелый труд провести анатомический анализ разрушений мозга, которые произвели он и Бробек.
И вот новый сюрприз. Стереотаксический прибор произвел разрушение совсем не в том месте, куда он был направлен, а именно он разрушил не амигдалу, а образование,.
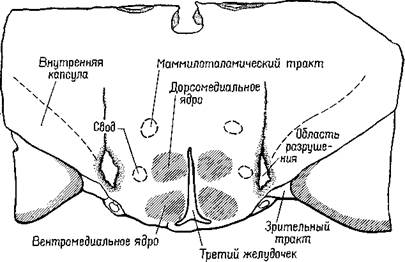
Рис. Х-4. Схема среза, проходящего через гипоталамус крысы, дающая представление о локализации в латеральном гипоталамусе «центра голода». Черным показаны области разрушения (Anand and Brobeck,
1951).
расположенное гораздо ближе к средней линии мозга — между амигдалой и гипоталамусом (рис. Х-4).
Таким образом, в латеральном гипоталамусе был открыт механизм голода, или центр Ананда — Бробека (Anand and Brobeck, 1955). Ананд, на протяжении ряда лет проводивший исследование этого вопроса, показал, что механизмы, побуждающие к еде, и механизмы насыщения находятся в реципрокных отношениях: когда центр насыщения при регистрации с помощью микроэлектродов обнаруживает активность, область, побуждающая к еде, тормозится. Когда же механизм насыщения находится в состояни покоя (как в случае низкого уровня сахара в крови), центр голода растормаживается и становится активным (Anand, 1963; рис. Х-5).
Оказалось, что центр голода и жажды в латеральном гипоталамусе состоит не из массы клеток, рецепторов, чувствительных к физико-химическому раздражению. Скорее, он представляет собой место пересечения путей от различных частей мозга, связывающих периферические и центральные станции, имеющие отношение к началу и прекращению еды и питья. Детали организации этой системы еще пока не установлены. Как уже указывалось, многое известно о взаимоотношении между ощущением голода и жажды и такими процессами, как сокращение и наполнение желудка, сухость во рту и т. д., которые составляют
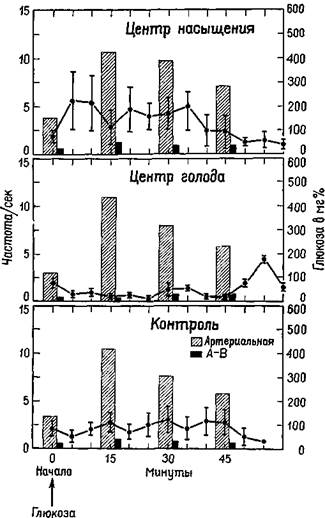
Рис. Х-5. График отдельно показывает среднюю частоту, нейронных разрядов, записанных от центра насыщения к центра голода, а также от контрольных областей. На рисунке представлена корреляция нейронных разрядов с уровнем глюкозы в артериальной крови и количеством усвоенной глюкозы (различие между артериальной и венозной концентрацией, А—В). По-видимому, изменение частоты нейронных разрядов от центра насыщения и центра голода лучше коррелирует с количеством усвоенной глюкозы, чем с ее абсолютным уровнем в артериальной; крови (см. Anand, 1963).
тшриферическую часть системы. Значительно меньше известно о центральных механизмах этой системы. Но начало уже положено (Brobeck, 1963).
НЕКОТОРЫЕ ФАКТЫ
Амигдала, с изучения которой начались наши эксперименты, является одной из структур, откуда идут волокна в латеральную гипоталамическую область пересечения нервных путей. Ее функции в регуляции питания и питья продолжают тщательно изучаться. Хотя инъекции малейших доз некоторых химических агентов (оказывающих адренэргическое или холинэргическое действие) в гипоталамический центр насыщения могут вызвать или остановить еду и питье (в зависимости от характера химического агента), такие же инъекции в амигдалу никакого эффекта не вызывают. Но если еда или питье уже начались, введение тех же самых химических агентов может изменить количество съеденной пищи или выпитой воды (Grossman, 1966; рис. Х-бб). Были установлены ясные количественные соотношения между дозой вводимого вещества и количеством поглощенной воды (Russel et al., 1968; рис. Х-ба), а кривая, характеризующая эти соотношения, напоминает многие кривые порогов, установленные в психофизических экспериментах, проведенных со зрительной и слуховой модальностями. Таким образом, психологические процессы экстероцептивных и висцеральных ощущений имеют много сходного.
Это совпадение, вероятно, не случайно. Однажды мне довелось обследовать некоторых больных с двусторонним удалением медиальной части височного полюса, включая амигдалу. Эти больные, так же как их двойники среди обезьян, ели значительно больше, чем нормальные люди, и их вес превышал нормальный на сто фунтов. Наконец, я спросил одного такого больного, как же он себя чувствует, постоянно ощущая голод. Но к моему большому удивлению, ожидаемого ответа не последовало. Одна больная, которая после операции прибавила за год в весе более чем сто фунтов (45 кг), была обследована во время завтрака. Была ли она голодна? Она ответила: «Нет». Хотела ли бы юна съесть кусок мяса, сочный бифштекс? Она ответила: «Нет». А хотела ли бы она шоколадную конфету? Она хмыкнула, но, когда конфету ей не дали, больше к этому предмету не возвращалась. Несколько минут спустя, когда обследование закончилось и открылись двери общей комнаты, она увидела других больных, завтракавших за длинным столом. Больная бросилась к столу, растолкала других больных и начала набивать едой рот обеими руками. Ее немедленно позвали в кабинет и повторили вопросы, относящиеся к еде. Были получены те же самые отрицательные ответы даже после многозначительного противопоставления этих
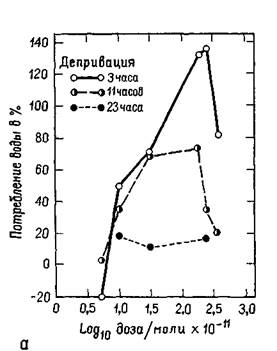
Рис. Х-6. А — отношения между изменением потребления воды и дозой карбахола (холин-эргетика), вводимого в амигдалу при трех уровнях водной депривации. Чтобы избежать отрицательных логарифмов, шкала по абсциссе представлена в молях ХЮ~" (RusselJ et ai., 1968);
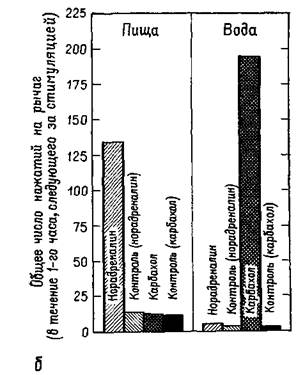
Б — влияние адренэргической и холинэргической стимуляции гипоталамуса у сытых крыс на скорость нажатий на педаль для получения пищевого и водного подкрепления по программе с варьирующим около 30 сек интервалом.
Контрольные уровни определялись в течение одночасового периода перед каждой стимуляцией (см, Grossman^
1962^.
1 4 16 64 256
Число нажатий на рычаг за кажЗую пищбую пилюлю
Рис. Х-7. Среднее число нажатий на рычаг (за каждые 12 чалов) у нормальных, ожиревших (поздняя стадия гиперфагии) и динамичных (до ожирения, ранняя стадия гиперфагии) животных в зависимости от числа нажатий, необходимых для получения пищевого шарика (Teitelbaum, 1957).
ответов ее недавнему поведению за столом. Повреждение мозга каким-то образом ослабило у больной ощущение голода и насыщения, и это ослабление сопровождалось чрезмерной едой.
Пока мы плохо понимаем, как происходит такое нарушение. Тем не менее этот пример ясно показывает, как нелепо предполагать, что между определенным типом поведения и инстроспек-тивным отчетом о нем существует прямое соответствие. Должны ли мы сказать, что больная ощущала голод, потому что она жадно ела, несмотря на ее отрицание наличия голода? Или мы должны рассматривать ее утверждения как истинные и искать объяснение ее неумеренной еды в чем-то другом? Этот парадокс может быть разрешен, если, как и в предыдущих главах, посвященных экстероцептивным ощущениям, мы будем считать, что поведение складывается из нескольких процессов, одним из которых является ощущение от своего состояния, описываемое в отчете испытуемого.
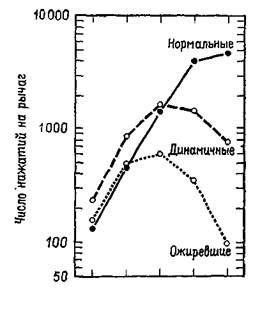
Рис. Х-8. Механизмы «остановки» и «запуска» (Stop and Go) в терминах схемы Т—О —Т —Е.
Аналогичный парадокс возникал перед исследователями функций гипоталамуса. Выше уже отмечалось, что, когда разрушения производились в области вентромедиального ядра гипоталамуса, оперированные крысы ели значительно больше контрольных и толстели. Но это еще не все. Хотя крысы с таким повреждением гипоталамуса ели больше, когда еда была вполне доступной, они в меньшей степени активно искали пищу, когда появлялись какие-либо препятствия (Miller, Bailey and Stevenson, 1950; см. также рис. Х-7). Было также обнаружено, что, чем вкуснее еда, тем больше ее съедают оперированные животные (Teitelbaum, 1955). Это наводит на мысль, что у оперированных животных больше не «влечение» к еде, а они более «разборчивы», нежели контрольные крысы. Результаты экспериментов, проведенных Кресном (1962) и Гроссманом (1966), усугубили парадокс: электрическое раздражение вентромедиального ядра вызывает прекращение поглощения пищи и воды у голодных крыс, а химическое раздражение холинэргического механизма вызывает топанье ногами (у песчанок, личное сообщение Глик-мана) и агрессивность, если животное рассердить (King and Hoe-bel, 1968). Гроссман коротко обобщил эти результаты в следующем утверждении: медиальный гипоталамус управляет изменениями висцеральных ощущений, а не потребностей. Но здесь мы снова сталкиваемся с нашей прежней дилеммой. Если медиальный гипоталамический механизм не связан с мотивацией, как же в таком случае возникает потребность в еде, питье и т. д.? Ответ
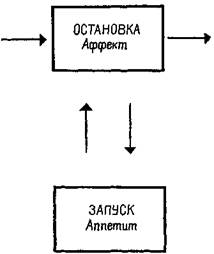
на этот вопрос дают эксперименты. Вентромедиальная и латеральная гипоталамические области составляют две части одной" системы: латеральная часть является механизмом, «запускающим» питание (удаление ее ведет у крыс к голоданию), а медиальная часть является тормозящим механизмом (рис. Х-8). Парадокс разрешается, если предположить, что процессы, которые выключают организм из активного состояния, также генерируют соответствующие ощущения. Таким образом, становится яснее важнейшее различие между мотивацией и эмоцией: термин «мотивация» можно относить только к потребностям, «запускающим» процессам (тем, которые объединяются в латеральной гипоталамической области), а термин «эмоция» — к действию аффективных, «останавливающих» или насыщающих процессов, восстанавливающих равновесие.
Многие из этих положений развиваются в следующей главе. Здесь же отметим только, что объяснения — те объяснения, с помощью которых ученые могут осмыслить полученные данные, — сформулированы в терминах изменения висцеральных ощущений, потребностей (мотивов) и аффектов (эмоций), получаемых в результате разрушения мозга. Выводы об этих изменениях в ощущениях мы делаем на основе наблюдаемых нейропси-хологических данных, но при объяснении экспериментальных результатов следует учитывать не только поведенческие, но и многие другие (например, нейрологические) факты.
РЕЗЮМЕ
Эксперименты, проведенные для уточнения функций рецеп-торных ядер мозга, привели к целому ряду загадочных парадоксов, которые не могли быть разрешены до тех пор, пока описание результатов осуществлялось на субъективном языке ощущений. Для каждой из рецепторных функций были установлены мотива-ционный, «запускающий», и эмоциональный, «останавливающий», механизмы. «Запускающие» процессы выражаются через элементарные потребности, а процессы «остановки» — через аффекты.
Г л а в а XI
ИНТЕРЕСЫ, МОТИВАЦИЯ И ЭМОЦИИ
МОДИФИКАТОРЫ ГОМЕОСТАТОВ
Фантастический успех и надежды, которые возлагались на нейрохимический подход к анализу многих висцеральных ощущений, составляющих «мир внутри нас», к сожалению, заслонили собой для большинства физиологически мыслящих исследователей некоторые другие проблемы, требовавшие разрешения. Именно этим проблемам и посвящена настоящая глава. В то время когда физиологическая наука была сосредоточена на изучении главным образом сенсорных и гуморальных механизмов, легко было утверждать, что восприятие является результатом переработки сенсорных процессов, а висцеральные ощущения — исключительно результатом действия гуморальных механизмов. К сожалению, такое представление продолжает существовать и в наше время, несмотря на то, что обилие новых нейропсихологи-ческих данных делает его сегодня несостоятельным.
В этом и состоит существо проблемы. Наблюдатели и экспериментаторы, работающие с людьми, неоднократно убеждались в недостаточности нейроэндокринного подхода при рассмотрении потребностей и чувств, субъективного опыта, всего того, что мы называем переживаниями.
Радость открытия, разочарование от неудачи, счастье понимания и тоска одиночества — эти чувства, по-видимому, далеки от уже описанных механизмов гомеостазиса.
Тип эксперимента, впервые проведенный С. Шехтером (Schachter and Singer, 1962), помогает подойти к только что обозначенной проблеме. Четыре группы студентов держали экзамен. Предварительно с этими студентами проводили эксперимент, в котором две группы экзаменовались в условиях враждебности, а две другие — в условиях дружественного отношения. На экзамене одна из каждой пары групп получила инъекцию адреналина, а другая — контрольную инъекцию физиологического раствора. Студенты давали отчет о своих переживаниях. Как и ожидалось,
первая группа испытывала преимущественно отрицательные эмоции, а вторая — положительные. Влияние же адреналина оказалось неожиданным. Он усиливал как положительные, так и отрицательные эмоции. Какое бы физиологическое состояние ни вызывала инъекция, его знак определялся установкой — социальным окружением студентов, а не введенным веществом.
Недавние экспериментальные результаты Шехтера (Schach-ter, 1968) подтверждают более ранние. Механизм регуляции голода (как он проявляется в еде) сейчас тщательно изучается. Шехтер считает, что, когда будет съедаться пища, определяют два класса переменных — внутренние и внешние. В большинстве случаев поглощение пищи имеет место тогда, когда внутренняя потребность и внешние возможности совпадают. С помощью своей, как всегда, остроумной (а иногда и несколько жестокой) методики Шехтер отделил случаи, когда главную роль играли ней-рогуморальные стимулы, от тех, когда определяющим была возможность поглощать пищу. Он показал, что тучный человек редко способен пренебречь возможностью поесть безотносительно к тому, каково его гуморальное состояние. Например, во время проведения эксперимента, в котором испытуемые отвечали на заданные им вопросы, им предлагали тарелку с печеньем. Разумеется, вопросы и ответы не имели отношения к самому эксперименту—экспериментатора интересовало только количество съеденного печенья. Обычная реакция среднего человека заключалась в том, чтобы отказаться от пищи, если «еще не время для еды».
«Еще не время для еды». Эта фраза содержит в себе первую проблему, которую я хотел бы подчеркнуть. Вы готовы есть, вы расположены есть благодаря действию гомеостатического механизма. Но этот механизм настраивается с помощью прошлого опыта; его отклонения регулируются вокруг «рабочей точки», установленной на основе жизненного опыта, во многом так же, как «поведение» парового котла определяется регулировкой термостата. Возвращаясь домой после теннисного матча, вы часто испытываете жажду, и глоток воды является как раз тем количеством, которое необходимо, чтобы утолить ее. Вы прекращаете пить задолго до того, как наступает требуемое для утоления жажды разжижение крови. Эксперименты, проведенные на собаках, показали, что такое поглощение воды глотками тонко «регулируется» потребностью в воде (Adolph, 1950; рис. XI-1). Работа системы, несомненно, меняется под влиянием прошлого опыта, даже когда система регулируется посредством гуморального механизма. У тучных людей такого рода явления также имеют место, но вместо гуморальных стимулов мотивацию создают вкусовые, обонятельные и даже зрительные стимулы. Для того чтобы вы не думали, что мотивация тучного человека совершенно отлична от вашей или моей мотивации нормального человека, со-
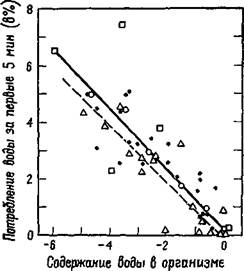
Рис. Х1-1. Типичные результаты эксперимента, обсуждаемого в тексте, которые показывают, что при жажде количество выпиваемой воды, когда для этого предоставляется возможность, пропорционально степени обезвоживания организма. Данные по отдельным собакам представлены разными символами (Adolph, 1943).
вершите путешествие (желательно без ограничения бюджета) и посетите пивные бары Мюнхена (особенно во время праздничных дней), венские кафе и рестораны Прованса. Затем, если вы отважитесь, измерьте свой вес по возвращении, особенно если вам эа 65 лет. Вы увидите, что основные гомеостаты мозга, испытавшие влияние прошлого опыта, находятся под контролем не только гуморальных стимулов, но и условий среды.
Внимание исследователей поведения животных было сосредоточено также на проблеме сенсорного контроля над мотивами и эмоциями. Существование множества названий для понятия влечения (drive), из которых наиболее распространенными являются «любознательность» и «побуждение», свидетельствует о том, что необходима более широкая база данных (помимо нейрогумораль-ных) для понимания «мотивации и эмоций».
«Время для...»—это то, что может быть включено в эту систему данных. В результате прошлого опыта внутри организма формируется набор установок, нервных моделей событий, пережитых в прошлом. До недавнего времени такой термин, как «установка», или «ожидание», не имел достаточного числа неопровержимых неврологических подтверждений. Это положение ради-
кально изменилось после классических опытов Е. Н. Соколова. Он показал, что всякий раз, когда изменяется конфигурация стимула, повторно воздействующего на сенсорный вход, по какому-либо из его параметров, происходит растормаживание и возникает ориентировочная реакция. Для изучения неврологии новизны (ориентировочной реакции и ее растормаживания) и привыкания были широко использованы поведенческие ответы (ориентировочные движения глаз, головы и тела), нейроэлектрические (ЭЭГ, низковольтная активность в изокортексе и тета-активность в гиппокам-пе) и висцеральные реакции (КГР, изменение частоты сердцебие1 ния и дыхания). Эти исследования говорят о том, что мозгу присущи некоторые модификации сервомеханизмов, такие, как гомеостаты и логические элементы типа Т—О—Т—Е в сенсорных системах.
Оказалось, что целая категория ощущений связана с более или менее гармоничными взаимодействиями, которые возникают между сервомеханизмами просто потому, что все они имеют место внутри одного и того же мозга. В гл. VII и VIII мы касались того, каким образом устанавливаются эти взаимоотношения, составляющие содержание гл. XIV, XV и XVI. Поэтому нам необходимо отметить только тот факт, что такие взаимодействия возникают благодаря организации основных элементов системы во временные микроструктуры, которые характеризуются определенными конфигурациями медленных потенциалов соединений, участвующих в этом процессе. Стабильность — лабильность такой общей организации является параметром, который можно изучать и который действительно широко изучается в понятиях ориентировочной реакции и процесса привыкания (Lacey and Lacey, 1958; Lacey et al., 1963). Мы часто сталкиваемся с ним в повседневной жизни, когда характеризуем личность как стойкую или нестойкую или когда признаемся, что пребываем в подавленном настроении.
АКТИВАЦИЯ КАК РЕАКЦИЯ НА НЕОПРЕДЕЛЕННОСТЬ
Что порождает эту неуравновешенность, это подавленное состояние духа? Факты говорят о том, что даже процесс зарождения таких состояний не прост.
Переживания «интересов», мотивов (потребностей) и эмоций (аффектов) являются результатом состояний, возникающих, когда организм сталкивается с новизной — новизной, которая создается вследствие того, что непрерывно меняющийся «мир внутри нас» приходит в соприкосновение со всегда изменяющимся «миром вне нас». В результате исследований, проведен-вых в моей лаборатории с использованием техники разрушения отдельных участков мозга, удалось выделить по крайней мере
два компонента ориентировочной реакции: один из них указывает на процесс исследования и опробования, а другой выявляется, когда организм фиксирует новизну. Только после такой фиксации возникает привыкание.
Результаты этих исследований были получены примерно в следующей последовательности. Несколько лет назад мы показали, что изменения темперамента и личности, легко вызываемые удалением височных долей мозга, связаны с удалением амигда-лы; эта структура височной доли, регулирующая потребности, обсуждалась в предыдущей главе (Pribram and Bagshaw, 1953; см. рис. Х-3, стр. 215). Дальнейший анализ показал, что амиг-дала участвует в различных типах поведения, которые мы обозначили как четыре фактора, развивая идеи Кеннона о реакциях «агрессии и бегства», возникающих при электрической стимуляции участков гипоталамуса (Pribram, 19606). Наши четыре фактора, дополняющие классификации Кеннона, включают пищевое и половое поведение. Поскольку между амигдалой и гипотала-мическими структурами существуют тесные анатомические связи, эти результаты казались вполне приемлемыми до тех пор, пока я не почувствовал недостаточность описательной корреляции между анатомией мозга и поведением и не попытался понять механизм этих отношений.
Если бы я удовлетворился тем, чтобы изучать поведение, как таковое, я, как и другие исследователи, поставил бы перед собой вопрос, не реализуется ли пищевое поведение, агрессия, бегство и половое поведение различными участками амигдалы. Когда были проведены эксперименты (с удалением или стимуляцией), направленные на разрешение этого вопроса, был получен, в сущности, отрицательный ответ. Но то, что я хотел узнать, касалось психологического процесса, того общего, что характеризовало эти виды аффективного поведения, поскольку одиночное разрушение (даже какой-нибудь целой анатомической формации) могло бы изменить одним ударом все эти различные типы поведения. Концепция «инстинкта» (Beach, 1955), несмотря на ее правдоподобность, по многим причинам была неудовлетворительной. Например, она не может объяснить, почему решающее влияние на изменение агрессивного поведения после удаления амигдалы оказывают социальные факторы (рис. XI-2). Это могут объяснить различные формы концепции «влечения» вследствие наличия сильных связей между механизмами амигдалы и гипоталамуса. Однако, как отмечалось ранее, такие концепции «влечения» также не в состоянии объяснить влияние разрушения и стимуляции гипоталамуса. Поэтому я решил избрать противоположный подход к проблеме и поставить вопрос следующим образом: будет ли поведение, которое явно не является врожденным или управляемым влечениями, испытывать на себе влияние удаления амигдалы?
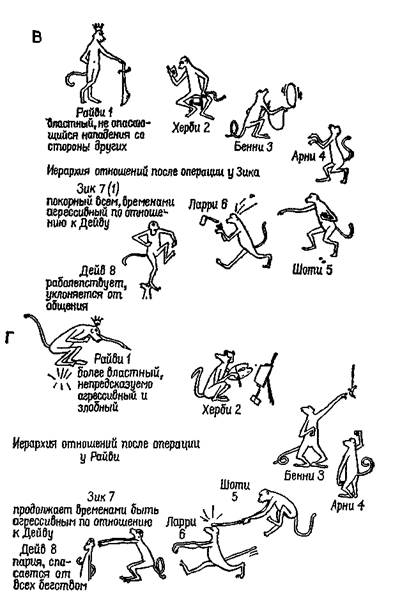
Рис. XI-2. А —иерархия господства в колония из восьми обезьян (резус)-самцов предподросгкового возраста до какого-либо хирургического вмешательства в мозг животных; Б—иерархия господства после двустороннего удаления амигдалы у Дейва. Обратите внимание, что он переместился на самую низкую ступень иерархии; В — иерархия господства после двустороннего удаления амигдалы у Зика; Г — окончательная социальная иерархия, установившаяся после двустороннего удаления амигдалы у Дейва,
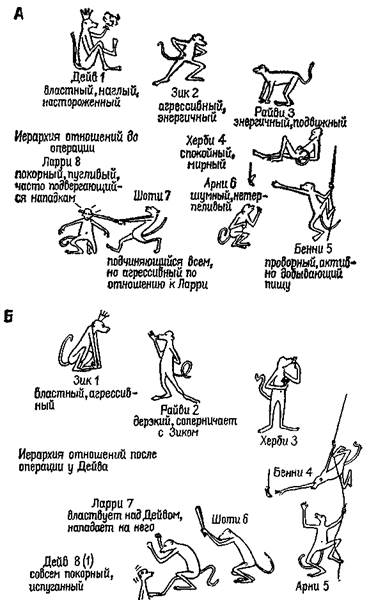
Зика и Райви. Следует отметить, что для Райви невозможно перемещение вниз по иерархической лестнице. Незначительные различия в размерах удаленного участка мозга у разных животных не могут объяснить различий в поведении, вызываемых этой операцией. Расхождение последних результатов с результатами предыдущих экспериментов обусловлено отсутствием агрессивности в характере Херби, занимавшего второе место на иерархической лестнице (Pribram, 1962).
Проведенные эксперименты не имели прямого отношения к исследованию упомянутых форм аффективного поведения. В сотрудничестве с Дж. Шварцбаумом (Schwartzbaum and Pribram, 1960), Мюриель Бэгшоу (Bagshaw and Pribram, 1965) и Э. Херс-том (Hearst and Pribram, 1964a, 19646) я провел эксперименты по переносу навыка. В одной серии экспериментов обезьян обучали выбирать из двух интенсивностей серого цвета более светлый. Затем во второй серии экспериментов, в которой проверялся перенос навыка, более светлая панель пары предъявлялась в сочетании с еще более светлой. Взятые для контроля нормальные обезьяны в соответствии с выработанным у них навыком продолжали выбирать из двух панелей более светлую. Обезьяны же с удаленной амигдалой во время тестовых испытаний действовали случайным образом, как если бы они решали совсем новую задачу (рис. XI-3). В другом эксперименте на перенос навыка обезьян ббучали выбирать из двух нарисованных квадратов квадрат большего размера, а затем проверяли, будут ли обезьяны выбирать большую из двух окружностей. Контрольные животные решали эту задачу, а животные с удаленной амигдалой — нет. Кроме того, в ситуации выработки оперантных условных реакций было проанализировано явление генерализации стимула. Обезьяны обучались регулировать освещение в экспериментальной камере, нажимая с разной частотой на педаль. Обезьяны с удалением амигдалы не отличались от контрольных и обнаружили такой же эффект генерализации, когда освещение в камере ступенчато менялось" по сравнению с тем, на которое они первоначально были натренированы. (Это поразительное отсутствие связи между переносом навыка и явлением генерализации уже само по себе интересно, но к рассматриваемому вопросу не имеет отношения.)
При выборе этих задач мы исходили из предположения, что их решение не требует участия гипоталамуса. Поскольку в обоих экспериментах при удалении амигдалы перенос навыка нарушался, один из моих выводов состоял в том, что процесс, контролируемый амигдалой, отличен от тех, которые обычно связывают с функцией гипоталамуса.
Ключом к разгадке того, что же представляет собой этот процесс, могут служить наблюдения за обезьянами, выполняющими задание с переменой стимулов. Как отмечалось ранее, обезьяны, лишенные амигдалы, не способны ни к переносу навыка, ни к выбору стимула по абсолютной величине. Во время тестовых экспериментов они вели себя, как в совершенно новой ситуации, и действовали на начальной стадии наугад (Douglas, 1966; Schwartzbaum and Pribram, 1960).
Продолжая эти наблюдения (Bagshaw and Benzies, 1968; Bagshaw and Coppock, 1968; Bagshaw, Kimble and Pribram, 1965; Bagshaw and J. Pribram, 1968; Kimble, Bagshaw and Pribram,
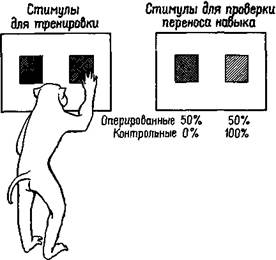
Рис. XI-3. Схема эксперимента для проверки переноса навыка у обезьян. После того как обезьяны были обучены выбирать из двух одновременно предъявляемых панелей более светлую (левый рисунок), им предъявлялся тест (правый рисунок) для проверки переноса навыка. В тестовом эксперименте более светлая панель оказывалась теперь более темной, что, естественно, создавало трудности для определения того, за какой панелью скрывается награда. Нормальные обезьяны продолжали выбирать более светлую панель независимо от ее абсолютной яркости. Обезьяны с удаленной амигдалой выбирали панель случайно, показывая тем самым, что они воспринимают тестовую ситуацию как «новую» и не связывают ее с предшествующим обучением (количественные данные см. на рис. XV-9).
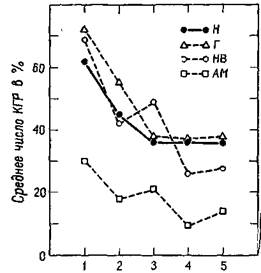
1965), мы показали, что в действительности при удалении амигдалы менялись реакции обезьян на новизну. Как по поведенческим, так и по электроэнцефалографическим показателям привыкание было длительным. С другой стороны, разрушение амигдалы вело к устранению висцеральных показателей ориентировочного рефлекса (КГР, изменение в частоте сердцебиения и дыхания) при отсутствии повреждения самих механизмов, ответственных за возникновение висцеральных реакций (рис. XI-4). Эти результаты говорят о том, что ориентировочный рефлекс на новизну проходит через две гипотетические стадии: первая, характеризующаяся поведенческими ориентировочными реакциями, «сравнивает», устанавливает новизну; вторая, связанная с
2J1
висцеральными реакциями, ведет к «фиксации» в нервной системе нового раздражителя и тем самым создает основу для привыкания (Pribram, 1969b). Таким образом, нарушения в поведении агрессии и бегства, пищевом и половом поведении, наблюдаемые в результате удаления амигдалы, я истолковал как результат тех затруднений, которые возникают в работе механизма, фиксирующего следы в нервной системе. Этот механизм необходим
'40
Серии по 10 проб
Рис. XI-4. Кривые, показывающие количество кожногальванических реакций — КГР (в %) за первые 50 предъявлений коротких тонов у нормальных обезьян (Н) и трех групп животных с двусторонним удалением гиппокампа (Г), нижневисочной коры (НВ) и амигдалы (AM) (Bagshaw, Kimble and Pribram, t965).
для организации поведения во времени. Проверка этой интерпретации предполагала, что поведение отсроченного чередования (выбор между двумя одинаковыми ящиками для пищи, которые заполнялись в последовательности справа — слева — справа — слева и т. д. при последовательных пробах) (Pribram, Lim, Poppen and. Bagshaw, 1966) также окажется нарушенным, и это предсказание подтвердилось.
Судя по этим результатам, амигдала выполняет гораздо большее число функций. Но это еще не все. Обращение к психологическому процессу фиксации помогло объяснить до сих пор не
объяснимое наблюдение (Miller, Galanter and Pribram, 1960, гл. 14). В гл. X говорилось о больной, у которой была произведена двусторонняя амигдалэктомия. За год после операции она сильно прибавила в весе. Казалось, что этот случай давал блестящую возможность выяснить непосредственно у больной, каковы были ее ощущения, заставлявшие ее есть так много. Она обычно отвечала, что ощущает очень немногое — она не чувствует, что очень голодна, то есть она не ощущала работу контролирующей и фиксирующей системы. Такая потеря регистрации — обычное явление в клинике у больных эпилепсией, возникающей вследствие повреждений вокруг амигдалы, повреждений, которые также порождают известные феномены deja vu (неадекватное переживание чего-то как уже знакомого) и jamais vu (неадекватное переживание известной ситуации как незнакомой). Таким образом, можно сказать, что контролирующие Образы характеризуют наши переживания, касающиеся «мира вне нас», во многом так же, как перцептивные образы характеризуют его свойства.
Эти эксперименты и их анализ позволяют подойти к рассмотрению второй, более важной и широко распространенной точки зрения на то, каким образом возникают переживания мотивов и эмоций, и прежде всего те переживания интереса, побуждений и аффектов, которые зависят от механизма активации (arousal). Только что упомянутые результаты экспериментов ставят вопрос о том, при каких условиях активация (измеряемая с помощью той же самой техники и тех же критериев, которые были использованы при исследовании амигдалы) приводит к фиксации и привыканию и при каких — к их нарушению. Классический ответ, который был дан на этот вопрос (Lindsley, 1951; Hebb, 1955), состоит в том, что уровень активации определяет его конечный эффект. Сейчас к этому можно добавить, что в результате изучения ориентировочных реакций стало известно, что уровень активации зависит от организации, конфигурации ожиданий, от состояния мозга, с которыми сопоставляется новый сигнал. Таким образом, уровень активации следует понимать как степень соответствия и несоответствия между конфигурациями, степень организации или дезорганизации, а не как величину возбуждения, которое меняется само по себе (см. Hebb, 1949; Лурия, 1960). С тех пор как для характеристики активации начали привлекать понятие степени организации, стало возможным выразить уровень активации через понятия информации и неопределенности. Количество информации или неопределенности выражается числом ответов «да» — «нет», необходимым для ответа на вопрос, являются ли два объекта воспринимаемого или переживаемого поля одинаковыми или различными. Неопределенность — обратная сторона информации; термин «неопределенность» употребляется как противоположный термину «информация», которая прямо пропорциональна определенному числу бит (двоичных
единиц «да» — «нет»). Чем больше это число, тем меньше неопределенность системы. Таким образом, можно было бы сказать, что такая точка зрения на теорию активации внесла в нее некоторую меру неопределенности.
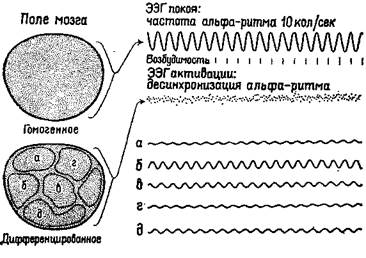
Само понятие «уровень» у сторонников теории активации, судя по их анализу, весьма неопределенно. Уровень активации — это не какое-то количественное изменение интенсивности, энергетического уровня центральной нервной системы, а нарушение равновесия, организации, измеряемой количеством неопределенности (и, следовательно, количеством информации), которое характеризует системы. На нейронном уровне активация сопровождается изменением динамических структур возбуждения в ретикулярной формации ствола мозга (см. Huttenlocher, 1961) и в коре (Burns, 1968, гл. 5), а не общего количества спайковых разрядов нейронов. Даже первоначальное определение активации как электроэнцефалографической реакции основано на том наблюдении, что в ЭЭГ возникает низковольтная высокочастотная активность и что такая активность указывает на десинхронизацию. Как было отмечено Линдсли (1961), это предполагает, что во время десинхронизации нервные элементы становятся функционально независимыми друг от друга и, таким образом, способными функционировать в качестве отдельных каналов, проводящих различную информацию. Эту возросшую сепарацию можно описать как увеличение степени организации или в терминах теории информации как увеличение неопределенности (рис. XI-5).
Естественным следствием такого понимания было бы предположение, что активация представляет собой состояние, при котором независимая активность нейронных ансамблей создает свободу для «резонанса» с другими, которые, несмотря на пространственную удаленность, имеют с ними общие характеристики. Таким образом, рассредоточенная информация в состоянии активации легче направляется по нужному адресу.
Гипотеза о том, что активация является в какой-то мере механизмом неопределенности, может показаться нам привлекательной. Но это еще не все, что известно в отношении активации. Например, в эксперименте, направленном на изучение того, какое влияние в ранний период жизни оказывает ограничение движений мы обнаружили, что инактивация младенца сопровождается увеличением лабильности автономной нервной системы (Konrad and Bagshaw, 1970).
Но прежде чем будут выяснены отношения между неопределенностью и контролем за поведением, нам необходимо еще раз вернуться к теории сервомеханизмов.
Результаты Шехтера еще не получили своего объяснения: усиления того или другого переживания в результате инъекции адреналина можно достигать путем количественного изменения состояния неопределенности, вызываемого введением определен-
Возбудимость!!» м ПН т П и ШИН «II НИ И III lit
Рис. XI-5. Схема гипотетических полей мозга и связанных с ними типов ЭЭГ. Альфа-ритм покоя на частоте 10/сек характеризует состояние гомогенности, или релаксации, мозга. Десинхронизированная, активированная ЭЭГ связана с переходом к дифференцированной структуре динамических полей мозга. Активированная ЭЭГ характерна для ситуации, требующей внимания и решения задач и вообще является характеристикой более эффективного восприятия и успешной деятельности. Исходя из того, что цикл возбудимости мозга связан с подъемом и падением амплитуды волн ЭЭГ, очевидно, что при ЭЭГ покоя сменяющие друг друга периоды возбудимости и невозбудимости могли бы возникать только с частотой 10 раз в сек. В случае же десинхронизированной ЭЭГ, возникающей в результате дифференциации мозгового ноля, периоды возбудимости следуют почти непрерывно (Linds-
1еу,(1961).
ного агента; специфичность же таких чувств, как враждебность или дружелюбие и т. д., еще требует объяснения.
КОРТИКАЛЬНЫЙ КОНТРОЛЬ И КИБЕРНЕТИКА ПРОЦЕССОВ УПРАВЛЕНИЯ
Специфичность чувств предполагает, что они являются скорее мотивами и эмоциями, чем просто количественным нарушением равновесия и неопределенностью или только нарушением протекания психических процессов. И конечно, в их основе лежит работа более сложной системы, чем гипоталамический механизм. Разнообразие переживаний интереса свидетельствует о том, что в устранении нарушений равновесия, в установлении специфических типов контроля, имеющих дело со специфическими ожида-
ниями (см. Zimbardo, 1969), участвует множество различных процессов.
Единственная возможность у организма управлять собой — это что-то делать, воздействовать на свое окружение. Как будет подробно описано в гл. XVI, всякий раз, когда сервосистема приобретает стабильность, появляются новые чувствительные индикаторы ее состояний и вводятся новые средства для управления ими. Например, когда термостаты были впервые установлены в домах, жители вначале столкнулись с более точными показателями снижения температуры: остывание наружных стен с закатом солнца вызывало поглощение тепла этими стенами. Поэтому были установлены термостаты для наружных стен, которые обеспечили более многосторонний контроль за отоплением в домах. Тот факт, что механизмы контроля развиваются как бы по спирали, обычно упускается из вида в самых распространенных определениях биологического гомеостатического процесса, что дает основание для возражений, выдвигаемых против биологической теории сервомеханизмов.
Но организм может достигать различных форм контроля не только посредством действия. Он может управлять, осуществляя самоконтроль, то есть он может создавать на основе своей неврологической системы внутренние регуляторы, — регуляторы, которые будут приводить к восстановлению равновесия даже и без внешнего действия. Положение, из которого мы исходим, заключается в том, что эти внутренние механизмы приспособления являются тем, что мы переживаем как эмоции.
Сейчас мы располагаем надежными нейрофизиологическими данными о том, что такие внутренние регуляторы не только возможны, но что указание на них стало уже общим местом. Было проведено множество экспериментов, чтобы показать, что входные каналы организма и даже сами рецепторы являются объектом контроля со стороны центральной нервной системы. Серии экспериментов, недавно выполненные в моих лабораториях, показали наличие кортикофугальных (кортико-подкорковых) влияний так называемых ассоциативных областей на периферию: на кохлеарное ядро и зрительный тракт (Dew-son, Nobel and Pribram, 1966; Nobel and Dewson, 1966; Spinelli and Pribram, 1966, 1967; Spinelli, Pribram and Weingarten, 1965; Spinelli and Weingarten, 1966; Weingarten and Spinelli, 1966). Было показано, что при стимуляции коры на этих уровнях нервной системы изменяется электрическая активность, вызванная щелчком и вспышкой света; в этих случаях изменяется как величина, так и форма рецептивных полей нейронов зрительной системы (см. рис. XI-6).
Данные об электрических ответах на вспышку света и щелчок представляют особый интерес. Когда реакция нервной системы вызывается двойным щелчком или двойной вспышкой
света, амплитуда ответа на второй стимул служит индикатором длительности, в течение которой часть системы занята обработкой первого входного сигнала. Подавление амплитуды второго ответа пары указывает таким образом на более длительный процесс восстановления — на большее время проведения сигнала по каналу. Такое увеличение времени свидетельствует о десинхрони-
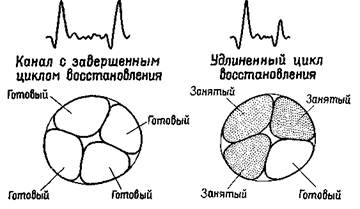
Рис. XI-6. Объяснение значения удлинения цикла восстановления возбудимости в афферентном канале после электрической стимуляции мозга. Эксперимент состоял в предъявлении пар стимулов, разделенных интервалами различной длительности. На график наносились изменения амплитуды ВП на второй стимул относительно амплитуды ВП на первый (в %) в зависимости от величины интервала, разделяющего эти стимулы (см. рис. XI-7). Предполагается, что, когда амплитуда ответа на второй стимул становится меньше амплитуды ответа на первый стимул, это означает, что афферентный канал в основном еще занят обработкой последствий действия первого стимула (показано на правой части рисунка). Обратите внимание на сходство этого рис. с рис. XI-5.
зации в канале, от которого зависит реакция на повторные воздействия: меньшее число волокон готово для проведения любого другого сигнала, предъявляемого в сериях. Удлинение цикла восстановления уменьшает, следовательно, избыточность, уменьшает число волокон, проводящих тот же самый сигнал по каналу. Таким образом, в любой момент через эту систему может быть проведено большее количество информации, большее число различных сигналов при условии, что система имеет достаточный резерв избыточности, чтобы препятствовать снижению эффективности, связанному с уровнем шума. Существует надежное доказательство, что это происходит именно так (Attneave, 1954; Barlow, 1961).
75 100 125 150
МштисекунЗы
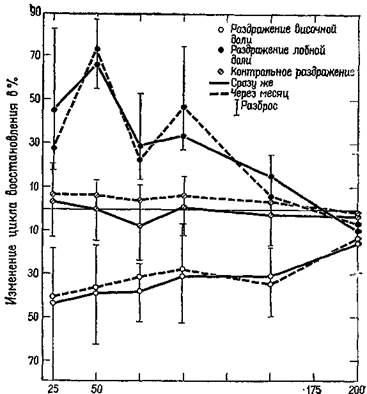
Рис. XI-7. Изменение цикла восстановления ответа афферентного канала под влиянием кортикальной стимуляции {рис. XI-6). Кора в течение нескольких месяцев подвергалась непрерывной электрической стимуляции (8 гц). Для контрольной стимуляции была использована теменная кора. Записи B1I производились немедленно после начала стимуляции и с этого времени повторялись еженедельно. На графике показаны функции восстановления при первых записях и через месяц. Вертикальные полосы показывают фактический разброс данных, полученных для каждой группы из четырех обезьян (Spinelli and Pribram, 1967).
Наши эксперименты показали далее, что стимуляция определенных участков коры височной доли (не амигдалы, а нижневисочной области) уменьшает избыточность, тогда как стимуляция другого участка коры (лобной) увеличивает избыточность в зрительной системе (рис. XI-7). Эти противоположные эффекты, по существу, либо «открывают» организм для воздействий со стороны его окружения, позволяя проводить большее число
238-
различных сигналов, большее количество информации в любой момент времени, либо, наоборот, «закрывают» входные каналы, создавая ограниченную проводимость для более узкого набора различных сигналов, для меньшего количества информации.
Результаты этих экспериментов говорят о том, что организм располагает по крайней мере двумя способами внутренне регулировать, контролировать неопределенность, но ни один из них не ведет к полной устойчивости поведения. Одним способом является увеличение скорости, с которой до него доводится информация, другим — снижение этой скорости. Один механизм делает организм «открытым» для внешних воздействий, другой ограничивает их. Управление посредством такого внутреннего контроля может, таким образом, быть достигнуто по крайней мере двумя путями: либо путем усиленного контроля за внешним окружением, либо путем доведения числа внешних воздействий до минимума и перехода к внутренним формам регуляции.
ПЕРЕЖИВАНИЯ И ИХ ВЫРАЖЕНИЕ
Теперь мы должны попытаться сделать еще один шаг в объяснении мотивация и эмоции, проблемы, которая стоит перед новой неврологией. Возможно, легче всего прибегнуть к традиционному делению психологических процессов на три класса и задать вопрос, каково различие между восприятием, действием и мотивацией — эмоцией. Сейчас имеется достаточно данных о том, что, когда организм воспринимает что-либо, он формирует внутренний образ своего окружения. Миллер, Галантер и я в книге «Планы и структура поведения» (1960, русский перевод 1965) утверждали, что, когда организм действует, он тем самым осуществляет внешнее выражение своих планов, нервных программ (сложных кодов) своего мозга. Ранее Толмен (1932) и Скиннер (1938) высказали мысль, что действие должно определяться через его внешние реакции, что оперантные условные «ответы» являются как бы метками поведения, которые можно зарегистрировать с помощью специальной записи. Теперь эти внешние проявления можно рассматривать как выражение нейронного процесса, сложившегося на основе тех условий, которые вызывают торможение неадекватных ответов, и в соответствии с ожиданиями организма, возникшими в результате прошлого действия сигналов и подкрепляющих агентов.
Если рассматривать мотивацию и эмоции в соответствии с этой схемой, становится очевидным, что они выражают отношения между восприятием и действием.
По-видимому, эти отношения состоят в следующем: когда разнообразие поступающей информации в значительной мере
превосходит совокупность действий, которыми располагает организм, он чувствует себя «заинтересованным», у него возникают мотивы и он делает попытки расширить набор своих действий. Всякий раз, когда его попытка оканчивается неудачей, не подкрепляется, вызывает фрустрацию или прерывается, организм непременно переживает эмоции, или, иначе говоря, вступают в действие механизмы, управляющие саморегуляцией, самоконтролем. Более того, возможно, что эмоции возникают тогда, когда, судя по прошлому опыту, вероятность подкрепления выполненного действия расценивается как низкая.
Противоположная ситуация порождает относительно «спокойное» состояние, не вызывающее мотивации и эмоций. Если репертуар возможных действий превосходит разнообразие имеющихся восприятий, наблюдается довольно любопытный ход событий. Весь репертуар действий, которыми располагает организм, начинает все больше определять богатство той информации, которую он получает. Так происходит до тех пор, пока этот процесс не будет завершен и не произойдет обратное, пока внешняя среда не начнет определять поведение. Такая ситуация возникает только тогда, когда организм ограничивает поступающую информацию и становится «закрытой системой». Чтобы снова стать открытой системой, чтобы вызвать интерес, организм должен открыть доступ разнообразным влияниям.
Мы предполагаем, что обычно интересы, переживания мотивов (влечений) и эмоций (аффектов) возникают тогда, когда организм пытается распространить свой контроль за пределы того, что он воспринимает. Пока эта попытка кажется осуществимой, организм испытывает влияние мотиваций: когда же эта попытка кажется невыполнимой, появляются аффекты, которые можно рассматривать как механизмы саморегуляции, направленные либо на то, чтобы преодолеть состояние, не поддающееся контролю, либо на то, чтобы подготовиться к другой попытке. Мотивация и эмоция возникают одновременно. Но мотив предполагает действие, формирование его внешнего выражения, эмоция же — противоположное, то есть отказ от данного действия, уход от него. Мотивация и эмоция, действие и переживания, определяющие эффективность поведения и его аффективную окраску, являются полярными, контролирующими эффект, механизмами. Они вступают в действие тогда, когда организм воспринимает больше, чем он может выполнить.
Те термины, которыми мы обозначаем переживания, связанные с появлением интереса, могут служить как для анализа эмоций, так и для анализа мотивов, хотя некоторое различие сохраняется: так, «быть влюбленным» относят к эмоции, «любить» — к мотивации; страх как эмоция имеет свое дополнение в страхе как мотиве. Эмоции и мотивы обычно тонко разграничены. Но при слишком сильном пассивном или активном выра-
?40
жении заинтересованности может происходить нарушение приспособления: слишком сильная эмоция ведет к разрушению или негибкости, срыву поведения.
Наконец, несколько слов о поведенческом выражении эмоциональных переживаний.
Согласно моему анализу, выражением мотивов являются действия, тогда как выражение эмоций отражает процесс внутреннего контроля. Для окружающих (как и для самого организма) такие выражения служат коммуникативными сигналами, которые полезно учитывать. Эти аффективные сигналы указывают на заинтересованность организма, однако они предполагают, что действие в данный момент по той или другой причине невозможно или не дает нужного эффекта.
Представленная здесь точка зрения на мотивы и эмоции как на результат регулируемых корой состояний формирования Образа, которые осуществляют и контролируют отношения между системами гомеостатов и получаемой информацией, исходящей не только из внутренней среды, но и из внешнего мира, дает нам новый подход к пониманию ряда важных психологических явлений.
РЕЗЮМЕ
Эксперименты, проведенные на людях, показывают, что возникновение мотивов и эмоций связано не только с механизмами ствола мозга. Мотивы и эмоции, как и эмоциональная заинтересованность, возникают, как правило, всякий раз, когда где-либо в мозгу нарушаются отношения между организованными нейронными структурами. Кибернетическая теория мотивации и эмоций, основанная на нейрофизиологических экспериментах, показывает, что контроль над этими отношениями осуществляет кора мозга.
ЧАСТЬ ТРЕТЬЯ

