




Лабораторная работа № 3
Грибоподобные организмы. Отделы Plasmodiophoromycota и Oomycota
Цель работы: изучить особенности строения вегетативных и генеративных структур грибоподобных организмов.
Задачи:
1) изучить биологические особенности грибоподобных организмов и экологические условия заражения растений патогенными грибоподобными организмами;
2) познакомиться с жизненным циклом паразитического слизевика, сапролегнии, фитофторы, альбуго;
3) изучить строение полового и бесполого спороношений исследуемых видов патогенных грибов.
Теоретические сведения:
Отдел Plasmodiophoromycota.
Отдел включает виды, имеющие внутриклеточный плазмодий. Они отличаются сложным циклом развития. В составе данного отдела один класс Плазмодиофоромицеты (Plasmodiophoromycetes). Класс Плазмодиофоромицеты (Plasmodiophoromycetes).
Представители класса являются внутриклеточными паразитами.
Плазмодиофора капустная – Plasmodiophora brassicae.
Plasmodiophora brassicae (плазмодиофора капустная) поражает корни растений из семейства крестоцветные. Заболевание известно под названием «кила капусты». Пораженные корни растения имеют уродливый вид, который придают им объемные опухоли. Плазмодий паразита находится внутри клеток корня растения. Под влиянием паразита нарушается синтез фенольных и индольных соединений в клетках растений, что приводит к увеличению размеров клеток (гипертрофии) и усиленному их делению (гиперплазии). Впоследствии пораженные корни загнивают. В клетках корня находятся многочисленные споры. При заражении капустной рассады кочаны могут совсем не развиваться. У зараженных взрослых растений они недоразвиваются. В цикле развития различают несколько стадий, протекающих в корневых волосках, почве и клетках коры корня растения.
Задание 1. Познакомьтесь с биологическими особенностями паразитического слизевика Plasmodiophora brassicae.
1) Зарисуйте внешний вид пораженного растения (гербарный образец).
2) Рассмотреть пораженные плазмодиофорой корни капусты на фиксированном материале. Изготовить временный водный препарат. Для этого взять небольшой кусочек пораженного корня капусты, сделать через него тонкий поперечный срез и поместить его в воду или КОН на предметном стекле. При малом и большом увеличении микроскопа рассмотреть клетки корневой паренхимы с плазмодием и спорами паразита.
3) Изучите строение спор и плазмодия, в котором хорошо видны при большом увеличении микроскопа вакуоли, бесцветные мелкие зернышки, и капельки масла. Зарисуйте общий вид пораженных клеток корня капусты.
4) Составьте схему цикла развития плазмодиофоры (рис. 1).
5) Опишите экологические условия заражения растений плазмодиофорой капустной.
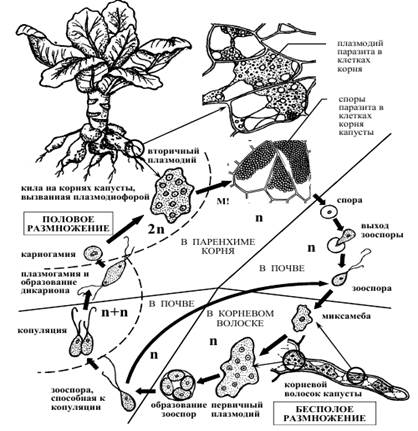
Рис. 1. Плазмодиофора капустная. Жизненный цикл.
Царство Страминопилы - Straminopila
Подцарство Гетеротрофные хромисты – грибоподобные организмы
Отдел Оомикота – Oomycota
Грибы, относящиеся к классу оомицетов, имеют хорошо развитый неклеточный мицелий, у немногих - ризомицелий. В клеточной стенке – целлюлоза. Митоз открытый, с центриолями. Кристы митохондрий трубчатые. Половой процесс – изогамия, или своеобразная оогамия, при которой содержимое антеридия не дифференцируется на гаметы. Бесполое размножение — зооспорами с двумя жгутиками (перистым и гладким). Отдел содержит классы: гифохитридиомицеты и оомицеты, В классе оомицеты несколько порядков, из которых главнейшие — сапролегниевые, пероноспоровые, лептомитовыеилагенидиевые (Г.А. Белякова, Ю.Т. Дьяков, К.Л. Тарасов Ботаника: водоросли и грибы, Т. 2, 2006, стр. 36-46). Водные грибы, в основном сапротрофы, реже паразиты.
Класс Оомицеты – Oomycetes
Порядок Сапролегниевые – Saprolegniales
Род Сапролегния – Saprolegnia.
Задание 2. Познакомьтесь с биологическими особенностями сапролегнии – Saprolegnia sp.
1) Изучите строение таллома и спороношений Saprolegnia sp.
Ход работы. Из сосуда с прудовой водой достаньте кусочек субстрата, на котором выращивалась Saprolegnia sp. (обычно её выращивают на белке сваренного вкрутую куриного яйца). Соскоблите с него белое опушение, которое и является талломом Saprolegnia sp.,перенесите на предметное стекло в каплю воды, накройте покровным стеклом. Приготовленный препарат рассмотрите сначала при малом (8 х 10), затем при большом (15 х 40) увеличении. При наличии зооспорангиев следует слегка постучать препаровальной иглой по покровному стеклу. При этом оболочка зооспорангиев лопается, а двужгутиковые зооспоры выплывают наружу.
Зарисуйте строение ценоцитного таллома Saprolegnia sp.
Обозначьте таллом, зооспорангии, зооспоры, жгутики зооспор, клеточную стенку таллома.
2) Отметьте особенности жизненного цикла сапролегнии (рис. 2).
Порядок Пероноспоровые – Peronosporales. Роды: Фитофтора – Phytophthora, Плазмопара – Plasmopara, Пероноспора – Peronospora.
Заражение фитофторой происходит путем попадания спор внутрь растения. Как правило, это происходит через самое незащищенное место — устьице (пора, на нижнем или верхнем слое эпидермиса листа, через нее происходит газообмен и испарение воды), для проникновения споры в устьице обязательно необходима вода. Зимуют споры фитофторы на зараженных клубнях, оставшихся в земле после сбора урожая, в почве, кучах ботвы или зараженных растениях. Болезнь распространяются от растения к растению, когда температура выше плюс 10°C, а влажность воздуха при этом сохраняется на уровне 75% в течение двух дней. Во время дождь споры смываются в почву, где происходит заражение молодых клубней. Также споры могут разноситься ветром на несколько километров от источника заражения.
Задание 3. Познакомьтесь с биологическими особенностями фитофторы Phytophthora infestans.
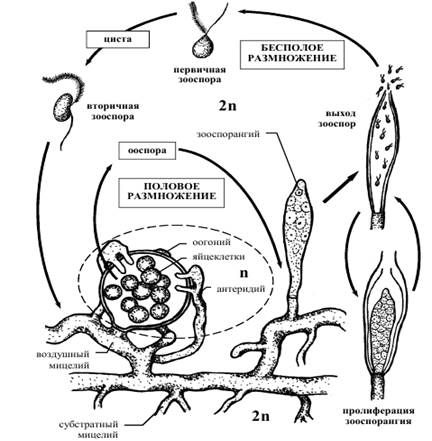
Рис. 2. Жизненный цикл сапролегнии
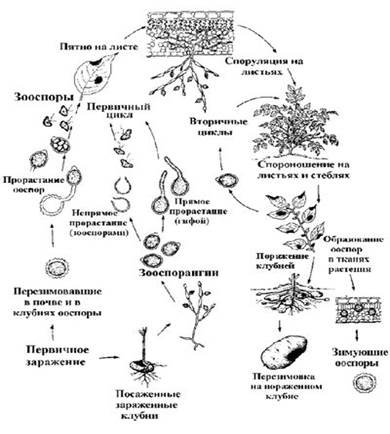
Рис. 3. Цикл развития фитофторы – Phytophthora infestans
1) Рассмотрите пораженные фитофторой ботву и клубни картофеля (гербарный образец).
2) Изготовьте временный водный препарат, соскоблив препаровальной иглой или лезвием немного налета с нижней стороны листа в каплю воды на предметное стекло, накрыть покровным стеклом и рассмотреть при большом увеличении микроскопа.
3) Изучите и зарисуйте конидиеносцы (спорангиеносцы) и конидии (спорангии) возбудителя фитофтороза.
4) Составьте схему жизненного цикла фитофторы (рис. 3), укажите условия заражения растений фитофторой.
Представители семейства альбуговые имеют хорошо развитый несептированный мицелий, распространяющийся в межклетниках растения-хозяина. У этих организмов наблюдается эволюция бесполых спороношений от типично спорангиальных, образующих зооспоры, к конидиальным, прорастающим гифой, что связано с переходом от водного к наземному образу жизни. Бесполое спороношение образуется в виде цепочек-зооспорангиев под эпидермисом растения-хозяина на булавовидных спорангиеносцах. Прорастают зооспорангии или зооспорами, или гифой; поэтому в некоторых руководствах обозначаются как конидии. Ооспоры – споры полового размножения – образуются в межклетниках растения-хозяина.
Задание 4. Изучите строение спороношений Albugo bliti.
1)От гербаризированного листа щирицы отделите небольшой фрагмент с выпуклым белым пятном – бесполым спороношением Albugo bliti – и размочите его в капле воды. Затем изготовьте срез, стараясь при этом, чтобы в область среза попал участок листа, на котором расположено спороношение. Полученные поперечные срезы листа поместите в каплю воды под покровное стекло и рассмотрите сначала при малом (8 х 10), затем – при большом (15 х 40) увеличении. На правильно приготовленном препарате будут видны цепочки зооспорангиев, выходящие наружу через разорвавшийся нижний эпидермис листа. В мезофилле листа хорошо заметны округлые бугорчатые ооспоры.
2) Зарисуйтестроение листа щирицы и спороношения Albugo bliti. Обозначьте верхний и нижний эпдермис листа, мезофилл, проводящий пучок (если он есть в пределах среза), ооспоры, зооспорангии, зооспоры.
Контрольные вопросы и задания:
1. Охарактеризуйте циклы развития плазмодиофоры капустной, вызывающей килу.
2. Каковы отличия оомицетов от других грибов?
2. Назовите особенности строения органов бесполого размножения оомицетов в связи с их образом жизни.
3. Охарактеризуйте циклы развития сапролегнии и фитофторы.
4. Что такое диморфизм и дипланетизм?
5. В чем сходство и отличие полового процесса в разных семействах оомицетов?
6. На примерах отдельных представителей оомицетов проследите путь постепенной приспособляемости грибов к наземным условиям существования, а также путь перехода грибов от сапротрофного к паразитическому образу жизни.
7. Назовите пероноспоровые грибы, являющиеся возбудителями опасных болезней культурных растений, и перечислите меры борьбы с ними.
Лабораторная работа № 4
Отделы хитридиомикотовые (Chytridiomycota) и зигомикотовые грибы (Zygomycota)
1. Общая характеристика настоящих грибов (Fungi)
2. Общая характеристика и основные представители хитридиомикотовых грибов (Chytridiomycota)
3. Отдел зигомикотовые грибы (Zygomycota), характерные черты строения и основные представители
1. Общая характеристика настоящих грибов (Fungi)
Первоначально единое царство грибы (Fungi) постепенно из одной системы к другой разделялось на ряд самостоятельных таксонов высшего порядка. Однако группа отделов в составе хитридиомикотовых (Chytridiomycota), зигомикотовых (Zygomycota), аскомикотовых (Ascomycota) и базидиомикотовых (Basidiomycota) грибов во всех системах оставалась единой, изменялся только ранг объединяющего их таксона высшего порядка. В учебном пособии Л. В. Гарибовой и С. Н. Лекомцевой (2005) – это царство грибы (Fungi). В системе И. Ю. Костикова с соавт. (2006), которого мы придерживаемся, подцарство настоящие грибы (Fungi) вместе с подцарствами растения (Plantae) и животные (Animalia) составляют царство пластинчатокристные (Platicristates).
У настоящих грибов имеются признаки, сближающие их с растениями: образование хорошо выраженной клеточной стенки; осмотрофное питание; размножение спорами; неподвижность в вегетативном состоянии; неограниченный рост; а также признаки, сближающие грибы с животными: первично гетеротрофный способ питания;
наличие в клеточной стенке хитина; запасной продукт – гликоген; выделение продукта жизнедеятельности – мочевины; отсутствие пластид и неспособность к фотосинтезу; путь синтеза лизина через аминоадипиновую кислоту (у растений – через диаминопимелиновую кислоту). Кроме того, имеются и специфические признаки: способность к синтезу маннитола (за исключением зигомикотовых грибов), собственные ферментативные ситемы синтеза триптофана, отсутствие комплекса Гольджи и его замена видоизменениями эндоплазматического ретикулума.
Вегетативное тело большинства грибов представляет собой мицелий, состоящий из ветвящихся нитей – гиф с апикальным (верхушечным) ростом и боковым ветвлением. Мицелий пронизывает субстрат и всей поверхностью поглощает из него питательные вещества (субстратный мицелий), а также располагается на его поверхности и может подниматься над субстратом (поверхностный и воздушный мицелий). На воздушном мицелии обычно образуются органы размножения.
Различают неклеточный (несептированный, или ценоцитический мицелий) лишенный перегородок и представляющий как бы одну гигантскую клетку с большим числом ядер, и клеточный (септированный) мицелий, разделенный перегородками – септами на отдельные клетки, содержащие от одного до многих ядер. Для представителей
отделов хитридиомикотовых и зигомикотовых характерен неклеточный мицелий. У представителей отделов аскомикотовых и базидиомикотовых мицелий клеточный, с настоящими септами, образующими упорядочение, синхронно делению ядер.
Септа развивается центрипетально, т. е. от стенки гифы к ее центру. В центре септы остается пора, через которую перемещаются питательные вещества, а также клеточные органеллы. Строение септы у разных групп грибов неодинаково. У аскомикотовых и некоторых базидиомикотовых образуется простая септа с порой в центре. Большинству базидиомикотовых свойственна так называемая долипоровая септа, имеющая трубчатое расширение у поры. С обеих сторон пора прикрыта мембранными структурами, называемыми парентосомами или поровыми колпачками.
У некоторых грибов, например дрожжей, вегетативное тело представлено одиночными почкующимися или делящимися клетками. Если такие почкующиеся клетки не расходятся, образуется псевдомицелий.
Некоторые примитивные грибы имеют одноклеточный таллом, иногда лишенный клеточной стенки. Такие одноклеточные талломы грибов способны развивать ризомицелий – разветвленные нитевидные структуры, лишенные ядер.
При формировании плодовых тел и некоторых вегетативных структур гифы грибов плотно переплетаются, образуя ложную ткань – плектенхиму. По происхождению она отличается от настоящей ткани, возникающей в результате деления клеток в трех направлениях.
Настоящие ткани у грибов встречаются довольно редко. Тканевое строение имеют, например, талломы лабульбениомицетов (Laboulbeniomycetes) из отдела аскомикотовые грибы.
У многих грибов гифы соединяются параллельно в мицелиальные тяжи, выполняющие проводящие функции. Хорошо развитые и дифференцированные мицелиальные тяжи называют ризоморфами. Они известны, например, у опенка осеннего. Ризоморфы достигают нескольких метров длины. Гифы их наружных слоев имеют утолщенные, часто темноокрашенные стенки и выполняют защитную функцию, а внутренние гифы – проводящую.
Другой тип видоизменений мицелия представляют широко распространенные у многих групп грибов склероции – плотные переплетения мицелия, служащие для перенесения неблагоприятных условий.
Клетки склероциев богаты запасными питательными веществами. Часто склероции дифференцированы на кору – наружные слои клеток, обычно толстостенных и темноокрашенных, и внутреннюю часть, состоящую из тонкостенных светлоокрашенных клеток.
Строение клетки. Клеточная стенка грибов содержит до 80–90 % полисахаридов, связанных с белками и липидами. Кроме того, в ее состав входят полифосфаты, пигменты, например меланины, и другие вещества. Микрофибриллярные скелетные компоненты клеточной стенки состоят из хитина. Аморфный матрикс образован преимущественно глюканами с разными типами связей.
Состав полисахаридов клеточной стенки специфичен для представителей крупных таксономических групп грибов. У хитридиомикотовых, аскомикотовых и базидиомикотовых грибов в клеточных стенках содержатся хитин и глюканы. По этому признаку зигомикотовые резко отличаются от перечисленных отделов: клеточные стенки
их мицелия содержат хитозан – полимер D-глюкозамина, но в них отсутствуют глюканы. У грибов с дрожжеподобным талломом, вне зависимости от их систематического положения количество хитина в клеточных стенках снижено, но содержатся маннаны – полимеры маннозы.
В цитоплазме клеток настоящих грибов хорошо различимы рибосомы, митохондрии и типичные эукариотические ядра. Протопласт грибов окружен цитоплазматической мембраной – плазмалеммой. На границе между цитоплазмой и вакуолью также расположена мембрана – тонопласт. Между тонопластом и плазмалеммой находится внутренняя мембранная система – эндоплазматическая сеть.
Митохондрии настоящих грибов имеют пластинчатые кристы и сходны с митохондриями высших растений и животных. По современным представлениям типичный аппарат Гольджи отсутствует, а его функцию выполняют различные видоизменения эндоплазматической сети. В растущих участках гиф в большом количестве содержатся пузырьки, или везикулы, происходящие от эндоплазматической сети. Они участвуют в транспорте веществ к месту синтеза клеточной стенки.
В цитоплазме клеток грибов часто присутствуют микротельца – круглые или овальные мембранные структуры. Возможно, они идентичны или являются предшественниками лизосом или пероксисом – органелл, содержащих соответственно гидролитические ферменты или каталазу.
В клетках грибов находится от одного до 20–30 ядер. Их размер обычно около 2–3 мкм. Они окружены оболочкой, состоящей из двух мембран. В нуклеоплазме содержатся ядрышко и хромосомы. При митотическом делении ядра ядерная оболочка часто сохраняется.
В клетках грибов присутствуют многочисленные включения: гранулы гликогена, капли липидов. В вакуолях часто находятся гранулы белков и волютина.
Подвижные клетки грибов – зооспоры и гаметы среди настоящих грибов характерны только для хитридиомикотовых. Жгутики настоящих грибов построены по типу, характерному для всех эукариот: их главный стержень (аксонема) состоит из двух одинарных центральных микротрубочек, окруженных кольцом из девяти дублетов микротрубочек. Аксонемы у грибов соединяются с кинетосомой, или базальным телом, разными способами. У некоторых хитридиомикотовых передняя часть ядра и его бока окружены так называемым ядерным колпачком, а кинетосома почти полностью опоясана большой полушаровидной митохондрией. Жгутики у настоящих грибов в отличие от псевдогрибов, бичевидные гладкие, расположены на заднем конце.
На поверхности одиночных клеток некоторых грибов – спор головневых и клеток дрожжей – обнаружены тонкие нитевидные структуры, названные фимбриями. Они имеют длину 0,5–10 мкм и толщину 6–7 нм. Предполагают, что фимбрии играют существенную роль в агрегации клеток.
Грибы размножаются вегетативным, бесполым и половым путем.
При вегетативном размножении от мицелия отделяются неспециализированные его части, которые дают начало новому мицелию. Одной из форм вегетативного размножения можно считать и образование хламидоспор – толстостенных клеток, предназначенных для перенесения неблагоприятных условий. У дрожжей вегетативное размножение происходит путем почкования клеток.
Бесполое размножение происходит при помощи специализированных неполовых клеток (реже многоклеточных структур) – спор. Споры у грибов развиваются эндогенно – внутри спорангиев или экзогенно – на особых веточках мицелия – конидиеносцах.
У зигомицетов в спорангиях развиваются неподвижные споры, называемые спорангиоспорами. Спорангии находятся на специализированных гифах – спорангиеносцах, поднимающихся над субстратом.
Конидии, как и спорангиоспоры, – неподвижные споры, пассивно переносимые токами воздуха, каплями дождя, насекомыми и другими способами. Однако в отличие от эндогенных спорангиоспор они образуются на мицелии или его специализированных ветвях – конидиеносцах экзогенно. Известно несколько типов формирования конидий из конидиогенных клеток. Конидиеносцы грибов имеют весьма разнообразное строение, часто наблюдается их агрегация в коремии, ложа или образование полузамкнутых структур – пикнид, в которых развиваются конидиеносцы и конидии. Бесполое размножение при помощи конидий характерно для базидиомикотовых и аскомикотовых, в том числе и анаморфных, где бесполое размножение достигло максимального разнообразия.
Половое размножение отмечено у всех групп грибов, кроме анаморфных, называемых также несовершенными грибами. Формы полового процесса у грибов разнообразны. Их можно разделить на три большие группы: гаметогамия, гаметангиогамия и соматогамия.
2. Общая характеристика и основные представители хитридиомикотовых грибов (Chytridiomycota)
Представители отдела хитридиомикотовые грибы (Chytridiomycota) еще тесно связаны с водной средой. Большинство паразитирует на водорослях, других водных грибах, водных высших растениях и на беспозвоночных животных. Некоторые встречаются как паразиты высших наземных растений, главным образом во влажной почве.
Значительно меньшая часть ведет сапротрофный образ жизни, поселяясь на ветвях, листьях, плодах, попавших в воду, на трупах насекомых и других животных, находящихся в воде. Немногие используют в качестве субстрата, а иногда и для питания сброшенные во время линьки или оставшиеся после гибели животного хитиновые покровы.
Вегетативное тело представителей этого класса у многих внутриклеточных паразитов в виде голой плазменной массы, очень похожей на плазмодий слизевиков. Оно всей своей поверхностью впитывает питательные вещества из клетки хозяина. Более высокоорганизованные формы имеют ризомицелий или зачаточный мицелий. У еще более высокоорганизованных паразитных хитридиомикотовых грибов вегетативное тело представлено ветвящимся ризомицелием, или системой гиф, который у паразитов может захватывать несколько клеток хозяина, а у сапротрофных грибов состоит из тонких ризоидов, внедряющихся в субстрат, и свободных ветвящихся гиф без перегородок. Основа клеточной стенки хитридиомикотовых грибов – хитин (до 60 %).
Бесполое размножение осуществляется зооспорами различного строения и формы с одним бичевидным гладким жгутиком, прикрепленным сзади. Зооспоры образуются в зооспорангиях. У некоторых хитридиомикотовых грибов все вегетативное тело превращается в спорангий, и жизнь данной особи прекращается (так называемые хо-локарпические формы), у других только какая-то часть становится зооспорангием, и зооспорангии могут развиваться по нескольку одновременно или последовательно (эукарпические формы).
Способы полового воспроизведения разнообразны. У некоторых в качестве гамет при определенных условиях функционируют зооспоры, у других сливаются сами особи (хологамия), у третьих развиваются одинаковые гаметы (изогамия) или различающиеся по размерам, а иногда, кроме того, по окраске и активности движения (гетерогамия). Наконец, есть формы с оогамным половым процессом. Гаметы образуются в специальных клетках – гаметангиях. Зигота превращается в покоящуюся клетку – цисту, одетую толстой оболочкой, содержащей хитин. В некоторых случаях цисты образуются без полового процесса. Большинство хитридиомикотовых грибов гаплоидны в вегетативном состоянии, и диплоидная фаза у них представлена зиготой, прорастающей после периода покоя, чаще всего как зооспорангии. У ряда видов имеются жизненные циклы со сменой поколений (иногда выделяется и дикарионтическая ядерная фаза).
Практически важен возбудитель «черной ножки» капустной рассады вид ольпидиум капустный (Оlpidium brassicae). Растения заражаются в парниках, особенно при избыточной влажности почвы и загущенных посевах, после появления на растениях семядолей или первых листьев. Стебель пораженного растения темнеет, утончается, нередко загнивает, а само растение поникает и гибнет.
Жизненный цикл. Заражение растения происходит зооспорами, которые при наличии воды прилипают к поверхности стебелька капусты, утрачивают жгутики и переливают свое содержимое в клетку эпидермиса (рис. 1).
Паразит не ограничивается клетками эпидермиса, а проникает в клетки первичной коры. Прорастание зооспоры сопровождается формированием многоядерной плазменной массы (плазмодий), которая покрывается оболочкой и превращается в зооспорангий с длинным горлышком, через которые зооспоры выходят наружу и поражают новые растения.
При неблагоприятных условиях зооспоры попарно копулируют, образую двужгутиковую зиготу, которая проникает в клетку хозяина, увеличивается, покрывается плотной бородавчатой оболочкой и превращается в цисту, которая является покоящейся стадией и зимует. Весной циста прорастает зооспорангием с зооспорами.
Очень важное практическое значение также имеет возбудитель рака картофеля вид синхитриум внутриклеточный (Synchytrium endobioticum) (рис. 2).
На клубнях картофеля, пораженного раком, появляются бугристые опухоли, напоминающие губку. Они разрастаются, часто превышая размеры клубня, чернеют и затем разрушаются. Вегетативное тело (плазмодий) желтое или красноватое, редко бесцветное.
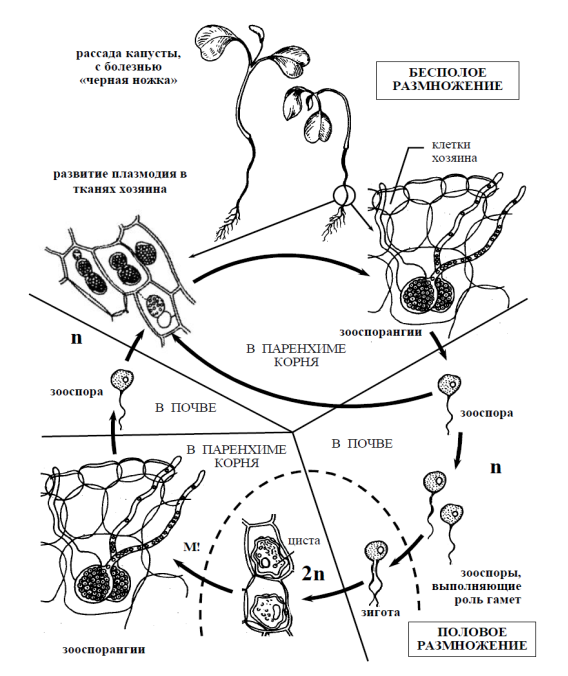
Рисунок 1. – Схема жизненного цикла ольпидиума капустного (Olpidium brassicae), возбудителя «черной ножки» рассады капусты
Жизненный цикл. Заражение клубней происходит так же, как у ольпидиума. Под действием паразита клетка эпидермиса разрастается, а соседние клетки усиленно делятся и оболочки их одревесневают.
Из таких клеток образуется розетка, в центре которой находится крупная клетка с телом паразита. Она одевается оболочкой и превращается в летнюю цисту, развивающуюся в сорус из 5–9 зооспорангиев, содержащих до 300 зооспор в каждом. Освобождаясь из разрушающейся опухоли, зооспоры снова заражают растения. Это может повторяться много раз в течение лета. Осенью в клубнях развиваются покоящиеся цисты (покоящиеся споры), которые могут сохраняться в почве много лет, и при благоприятных условиях прорастают пузыревидным выростом, в котором содержимое цисты распадается на отдельные зооспорангии.
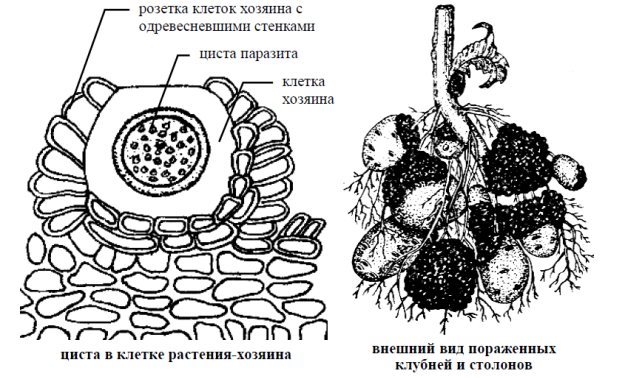 Рисунок 2. – Синхитриум внутриклеточный (Synchytrium endobioticum), возбудитель рака картофеля
Рисунок 2. – Синхитриум внутриклеточный (Synchytrium endobioticum), возбудитель рака картофеля
Заболевание встречается в Европе, США, Канаде, Южной Америке, Японии, Южной Африке. Потери урожая от рака могут составлять 40–60 %. Основные меры борьбы – выведение устойчивых сортов и обеззараживание почвы.

