




Цель:
· Умение регистрации электроэнцефалограммы и принципы анализа.
· Изучении внешнего электрического поля мозга при помощи ЭЭГ.
· Значение для генеза ЭЭГ взаимосвязи электрической активности пирамидных нейронов.
Основные вопросы темы:
1.Какие методы используется для регистраций ЭЭГ?
2.Основные типы электрической активности пирамидных нейронов.
3.Какие современные модели используется в ЭЭГ?
4.Какое значение имеет взаимосвязь электрической активности пирамидных нейронов.
5.Какое важное условие генеза ЭЭГ?
Методы обучения и преподования: Работа группах
Краткое содержание по теме
Исследование рабочих свойств центральной нервной системы производится при помощи специальных нейрофизиологических методов. Одним из основных является электроэнцефалография, позволяющая регистрировать суммарную активность нейронов коры головного мозга, представляющую собой колебательный процесс в частотном диапазоне в основном от 1 до 30-40 колебаний в секунду и регулирующуюся глубинными мозговыми структурами. Таким образом, по картине активности коры головного мозга возможно оценить и ее самое, и степень подкорковых влияний на процесс ее формирования.
Электроэнцефалография (ЭЭГ) (электро- + др.-греч. ενκεφαλος — "головной мозг" + γραφω — "пишу", изображать) — раздел электрофизиологии, изучающий закономерности суммарной электрической активности мозга, отводимой с поверхности кожи головы, а также метод записи таких потенциалов. Электроэнцефалография дает возможность качественного и количественного анализа функционального состояния головного мозга и его реакций при действии раздражителей. Запись ЭЭГ широко применяется в диагностической и лечебной работе (особенно часто при эпилепсии), в анестезиологии, а также при изучении деятельности мозга, связанной с реализацией таких функций, как восприятие, память, адаптация и т. д. Регистрация ЭЭГ осуществляется с помощью новейшего 32-канального электроэнцефалографа «Нейрон-Спектр-5» (рис-1). Многоканальная запись ЭЭГ позволяет одновременно регистрировать электрическую активность всей поверхности мозга, что дает возможность проводить самые тонкие исследования.
Достоинствами метода электроэнцефалографии являются объективность, воз-можность непосредственной регистрации показателей функционального состояния мозга, количественной оценки получаемых результатов, наблюдения в динамике. Большое преимущество этого метода состоит в том, что он не связан с вмешатель-ством в организм обследуемого.
Метод ЭЭГ является наиболее адекватным для изучения нейрофизиологиче-ских основ психической деятельности, оценки зрелости центральной нервной системы и общего функционального состояния мозга. Когерентный анализ ЭЭГ позволяет оценить степень согласованности электрической активности в разных точках головного мозга, что даёт возможность исследования особенностей функционирования мозга как единого целого.
ЭЭГ является клиническим методом исследования, позволяющим диагности-ровать эпилепсию, выявить возможные дегенеративные, опухолевые поражения головного мозга, установить их локализацию (рис.2).
 Начало изучению электрических процессов мозга было положено Д. Реймоном в 1849 году, который показал, что мозг, также как нерв и мышца, обладает электрогенными свойствами. Начало электроэнцефалографическим исследованиям положил В. В. Правдич-Неминский, опубликовав 1913 году первую электроэнцефалограмму записанную с мозга собаки. В своих исследованиях он использовал струнный гальванометр. Так же Правдич-Неминский вводит термин электроцереброграмма.
Начало изучению электрических процессов мозга было положено Д. Реймоном в 1849 году, который показал, что мозг, также как нерв и мышца, обладает электрогенными свойствами. Начало электроэнцефалографическим исследованиям положил В. В. Правдич-Неминский, опубликовав 1913 году первую электроэнцефалограмму записанную с мозга собаки. В своих исследованиях он использовал струнный гальванометр. Так же Правдич-Неминский вводит термин электроцереброграмма.
Рис. 1.
Первая запись ЭЭГ человека получена австрийским психиатром Гансом Бергером в 1928 году. Он же предложил запись биотоков мозга называть «электроэнцефалограмма». Работы Бергера, а также сам метод энцефалографии получили широкое признание лишь после того как в мае 1934 года Эдриан и Мэттьюс впервые убедительно продемонстрировали «ритм Бергера» на собрании Физиологического общества в Кембридже.
Регистрация ЭЭГ производится специальными электродами (наиболее распространенные мостиковые, чашечковые и игольчатые). В настоящее время чаще всего используется расположение электродов по международным системам «10—20 %» или «10-10 %». Каждый электрод подключен к усилителю. Для записи ЭЭГ может использоваться или бумажная лента или сигнал может преобразовываться с помощью АЦП и записываться в файл на компьютере. Наиболее распространена запись с частотой дискретизации 250 Гц. Запись потенциалов с каждого электрода осуществляется относительно нулевого потенциала референта, за который принимается мочка уха, или кончик носа. В настоящее время получают все большее распространение перерасчет потенциала относительно взвешенного среднего референта, за который принимается все каналы с определенными весовыми коэффициентами. При таком расчете возможные артефакты локализуются, а влияние соседних отведений друг на друга уменьшается.
Рис. 2.
Показания для ЭЭГ:
- черепно-мозговые травмы - для оценки функционального состояния мозга и судорожной готовности;
- проведение ЭЭГ в динамике для оценки эффективности противосудорожной терапии;
- синдром вегетативной дисфункции с паническими вегетативными пароксизмами;
- дифференциальная диагностика тсинкопальных состояний с целью исключения эпилептической активности.
В зависимости от частоты колебаний выделяется несколько ритмических рисунков электрической активности мозга – ритмов. Так, альфа–ритм, в большинстве случаев наиболее широко представленный в электроэнцефалограмме взрослого человека, имеет частотный диапазон от 8 до 13 колебаний в секунду и тесно связан в своем происхождении с системой зрительного восприятия. Поэтому он наиболее отчетлив он при закрытых глазах, то есть в состоянии ее максимального покоя, и лучше всего выражен в затылочных отделах, то есть там, где располагается высший отдел анализа зрительной информации. Наиболее высокочастотная часть электрической активности мозга, превышающая рамки альфа–ритма по частоте, именуется бета-активностью. Амплитуда ее, как правило, невысока и выражена она в противовес альфа-ритму, больше лобной и височной проекциях. Эта высокочастотная активность чаще всего рассматривается как признак активной работы многочисленных ансамблей нервных клеток. Альфа и бета активностью оканчивается ряд ритмических рисунков, характерных для взрослого человека в состоянии покоя, однако выделяются еще два варианта мозговой активности – тета и дельта. Тета-диапазон – более медленный в сравнении с альфа, от 7 до 5 колебаний в секунду. Дельта-волна еще медленнее, в секундном отрезке записи она может уместиться лишь 1-4 раза. Для такого рода медленной активности в состоянии бодрствования имеется в медицинской практике синоним – патологическая, то есть связанная с патологией, или – заболеванием, мозга. Ритмический рисунок мозговой активности существенно меняется с возрастом. Так, со второго полугодия сначала появляется, а затем постепенно начинает преобладать в картине активности альфа-ритм. Интересные метаморфозы происходят с медленной активностью. Патологической она считается только для взрослых в состоянии бодрствования. У детей наличие медленных волн в электроэнцефалограмме является нормальным, а вот представленность их отчетливо уменьшается с возрастомю. Большинство имеющихся экспериментальных данных говорит о том, что генез ЭЭГ определяется в основном электрической активностью коры больших полушарий головного мозга, а на уровне клеток – активностью ее пирамидных нейронов. У пирамидных нейронов выделяют два типа электрической активности. Импульсный разряд (потенциал действия) с длительностью около 1 мс и более медленное (градуальное) колебание мембранного потенциала – тормозные и возбуждающие постсинаптические потенциалы (ПСП). Тормозные ПСП пирамидных клеток генерируются в основном в теле нейрона, а возбуждающие ПСП – преимущественно в дендритах. Правда, на теле нейрона имеется определенное количество возбуждающих синапсов, и соответственно этому тело пирамидных нейронов(сома) способно генерировать также и возбуждающие ПСП. Длительность ПСП пирамидных клеток по крайней мере на порядок больше продолжительности импульсного разряда.
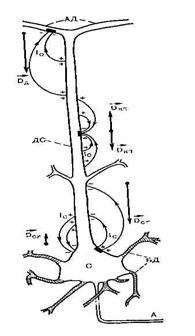
Изменение мембранного потенциала обусловливают возникновение в пирамидных клетках двух токовых диполей, отличающихся по цитологической локализации (рис3).
. Один из них – соматический диполь с дипольным моментом 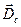 . Он формируется при изменении мембранного потенциала тела нейрона; ток в диполе и во внешней среде протекает между сомой и дендритным стволом. Вектор дипольного момента при импульсном разряде или генерации в теле нейрона возбуждающего ПСП направлен от сомы вдоль дендритного ствола, а тормозной ПСП создает соматический диполь с противоположным направлением дипольного момента. Другой диполь, называемый дендритным, возникает в результате генерации возбуждающих ПСП на ветвлении апикальных дендритов в первом, плексиморным слое коры; ток в этом дипооле течет между дендритным стволом и указанным ветвлением. Вектор дипольного момента
. Он формируется при изменении мембранного потенциала тела нейрона; ток в диполе и во внешней среде протекает между сомой и дендритным стволом. Вектор дипольного момента при импульсном разряде или генерации в теле нейрона возбуждающего ПСП направлен от сомы вдоль дендритного ствола, а тормозной ПСП создает соматический диполь с противоположным направлением дипольного момента. Другой диполь, называемый дендритным, возникает в результате генерации возбуждающих ПСП на ветвлении апикальных дендритов в первом, плексиморным слое коры; ток в этом дипооле течет между дендритным стволом и указанным ветвлением. Вектор дипольного момента 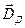 дендритного диполя имеет направление в сторону сомы вдоль дендритного ствола.
дендритного диполя имеет направление в сторону сомы вдоль дендритного ствола.
Генерация возбуждающего ПСП в районе дендритного ствола без ветвления приводит к появлению квадруполя, поскольку при этом от частично деполяризованного участка ток внутри клетки распространяется в двух противоположных направлениях, в результате чего формируются два диполя с противоположным напрвлением дипольных моментов 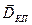 . Так как диполи малы по сравнению с расстояниями до точек отведения ЭЭГ, внешним полем квадрупольного генератора пирамидных клеток можно пренебречь.
. Так как диполи малы по сравнению с расстояниями до точек отведения ЭЭГ, внешним полем квадрупольного генератора пирамидных клеток можно пренебречь.
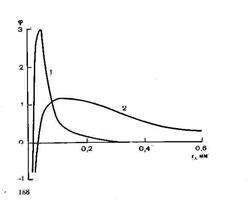 На (рис 4) изображена полученная пространственная структура электрического поля вдоль дендритного ствола и вокруг на расстоянии около 0,01 мм от продольной оси этого ствола. Оказалось, что внешнее поле пирамидного нейрона при импульсном разряде очень резко уменьшается вдоль дендритного ствола: уже на расстоянии около 0,3 мм потенциал падает практически до нуля. В противоположность этому внеклеточное ПСП характеризуется гораздо большей протяженностью (примерно на порядок), и, следовательно, при этой активности пирамидные клетки имеют гораздо более высокий дипольный момент. Это различие находит обьяснение при рассмотрении пассивных электрических свойств дендритного ствола.
На (рис 4) изображена полученная пространственная структура электрического поля вдоль дендритного ствола и вокруг на расстоянии около 0,01 мм от продольной оси этого ствола. Оказалось, что внешнее поле пирамидного нейрона при импульсном разряде очень резко уменьшается вдоль дендритного ствола: уже на расстоянии около 0,3 мм потенциал падает практически до нуля. В противоположность этому внеклеточное ПСП характеризуется гораздо большей протяженностью (примерно на порядок), и, следовательно, при этой активности пирамидные клетки имеют гораздо более высокий дипольный момент. Это различие находит обьяснение при рассмотрении пассивных электрических свойств дендритного ствола.
По отношению к потенциалу действия ввиду его кратковременности
Рис.3. мембрана дендрита ведет себя как емкость, обладающая низким сопротивлением току высокой частоты. Поэтому ток, обусловленный импульсной активностью, циркулирует на небольшом расстоянии от тела клетки; емкость мембраны шунтирует отдаленные участки ствола. Действительно, по данным микроэлектродных исследований, внешнее электрическое поле пирамидных нейронов, генерируемое потенциалом действия, не обнаруживается рис.4.
уже на расстояниях выше 0,1 мм. таким образом, ЭЭГ должна в основном создаваться «медленным» соматическим и дендритным диполями, возникающими при генерации тормозных и возбуждающих постсинаптических потенциалов.
При изучении внешнего электрического поля мозга регистрируют и интерпретируют переменный сигнал ЭЭГ, а постоянную составляющую, как правило, не принимают во внимание. Как видно на(рис. 5), ЭЭГ фоновой активности мозга представляет собой весьма сложную зависимость разнсти потенциалов от времени и выглядит как совокупность случайных колебаний разности потенциалов. Для характеристики таких хаотических колебаний («шумов») используют параметры, известные из теории вероятности: среднюю величину и стандартное отклонение 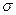 от средней величины. Чтобы наити , выделяют
от средней величины. Чтобы наити , выделяют
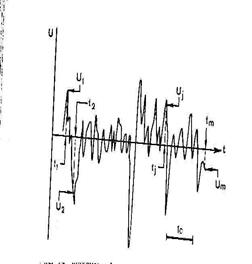 участок на ЭЭГ, который разбивают на небольшие равные интервалы времени, и в конце каждого интервала (ti, tj, tm на рис. 74) определяют напряжение U (Ui, Uj, Um на рис. 74). Стандартное отклонение рассчитывают по обычной формуле:
участок на ЭЭГ, который разбивают на небольшие равные интервалы времени, и в конце каждого интервала (ti, tj, tm на рис. 74) определяют напряжение U (Ui, Uj, Um на рис. 74). Стандартное отклонение рассчитывают по обычной формуле: 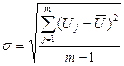 , (1.1)
, (1.1)
в которой 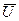 - среднеарифметическое значение разности потенциалов;
- среднеарифметическое значение разности потенциалов; 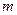 - число отсчетов
- число отсчетов 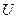 . При отведении ЭЭГ от твердой мозговой оболочки величина для фоновой активности составляет 50-100 мкВ.
. При отведении ЭЭГ от твердой мозговой оболочки величина для фоновой активности составляет 50-100 мкВ.
Аналогичная характеристика (стандартное
Рис.5. отклонение) используется и для описания градуальной активности отдельных нейронов 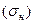 . При изучении ритмических ЭЭГ, характеризующихся определенной амплитудой и частотой изменение разности потенциала, показателем величины ЭЭГ может служить амплитуда этих колебаний.
. При изучении ритмических ЭЭГ, характеризующихся определенной амплитудой и частотой изменение разности потенциала, показателем величины ЭЭГ может служить амплитуда этих колебаний.
В настоящее время в исследованиях ЭЭГ для моделирования электрической активности коры головного мозга рассматривают поведение совокупности токовых электрических диполей отделных нейронов. Предложено несколько таких моделей, позволяющих объяснить отделные особенности ЭЭГ. Рассмотрим модель М. Н. Жадина, которая на примере генеза ЭЭГ при отведении с твердой мозговой оболочки позволяет выявить общие закономерности возникновения суммарного внешнего электрического поля коры.
Основные полежения модели: 1) внешнее поле головного мозга в некоторой точке регистрации – интегрированное поле, генерируемое токовыми диполями нейронов коры; 2) генез ЭЭГ обусловен градуальной электрической активностью пирамидных нейронов; 3) активность разных пирамидных нейронов в определенной степени взаимосвязана (скорелирована); 4) нейроны распределены по коре равномерно и их дипольные моменты перпендикулярны к поверхности коры; 5) кора плоская, имеет конечную толчину 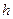 , а ее остальные размеры бесконечны; со стороны черепа мозг ограничен плоской бесконечной токонепроводящей средой. Обоснование первых двух положений расмотрено выше. Остановимся на друних положениях модели.
, а ее остальные размеры бесконечны; со стороны черепа мозг ограничен плоской бесконечной токонепроводящей средой. Обоснование первых двух положений расмотрено выше. Остановимся на друних положениях модели.
Очень большое значение для генеза ЭЭГ имеет взаимосвязь электрической активности пирамидных нейронов. Если бы градуальное изменение мембранного потенциала во времени происходило в каждом нейроне совершенно независимо от остальных клеток, переменная составляющая потенциала их суммарного внешнего электрического поля была бы неболшой, так как увеличение потенциала за счет усиления активности одного нейрона в значительной мере скомпенсировалось бы хаотическим снижением активности других нейронов. Сравнительно высокая величина регистрируемой в опыте ЭЭГ заставляет предположить, что между активностями пирамидных нейронов существует положительная кореляция. Количественно это явление характеризуют коэффициентом корреляции 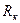 . Этот коэффициент равен нулю при отсутствии связи между активностями индвидуальных нейронов и был бы равен единице, если бы изменение мембранного потенциала (дипольных моментов) клеток происходили совершенно синхронно. Наблюдаемое в действительности промежуточное значение свидетельствует о том, что деятельность нейронов синхронизована лишь частично.
. Этот коэффициент равен нулю при отсутствии связи между активностями индвидуальных нейронов и был бы равен единице, если бы изменение мембранного потенциала (дипольных моментов) клеток происходили совершенно синхронно. Наблюдаемое в действительности промежуточное значение свидетельствует о том, что деятельность нейронов синхронизована лишь частично.
Интегрированное поле множества диполей-нейронов было бы очень слабым при высоком уровне синхронизации, если бы векторы дипольных моментов элементарных источников тока были ориентированы в коре хаотически. В этом случае наблюдалась бы значительная взаимная компенсация полей индивидуальных нейронов. В действительности же, согласно цитологическим данным, дендритные стволы пирамидных клеток в новой коре (эти клетки составляют 75% от всех клеток коры) ориентированы практически одинакова, перпендикулярно поверхности коры. Поля, создаваемые диполями таких одинакова ориентированных клеток, не компенсируются, а складываются. Расчеты, произведенные на оснований всех этих положений, показали, что для ЭЭГ, отводимой от твердой мозговой оболочки,
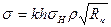 (1.2)
(1.2)
где k - коэффициент, численно равный средней плотности пирамидных нейронов в коре; ρ – удельное сопротивление коры: σH – среднее стандартное отклонение изменения во времени дипольного момента нейронов; Rk – средний попарный коэффициент корреляции активности нейронов. Из независимых опытов можно найти параметры, входящие в формулу (1.2). Так, для кролика h ≈ 0,0017 м, ρ ≈ 3 Ом ∙ м, k ≈ 4 · 1013. По расчетам А. М. Гутмана, σН ≈ 6 ∙ 10-15 А · м. Если принять Rk = 0,003, то рассчитанное отклонение ЭЭГ будет около 70 мкВ, т.е. очень близко к действительным величинам. Принятое значение коэффициента корреляции Rk (а иногда и вомного раз больше) наблюдается в опытах при исследовании различных типов электрической активности мозга.
Таким образом, внешнее электрическое поле головного мозга, обнаруживаемое в виде ЭЭГ, можно рассматривать как результат сложения полей пирамидных нейронов новой коры. Важное условие такого генеза ЭЭГ – одинаковая ориентация диполей нейронов и наличие положительной корреляции в их градуальной электрической активности; при этом достаточен даже очень низкий уровень попарной корреляции. Получить информацию о соответствии показателей работы мозга имеющимся нормативным данным. Поскольку запись активности производится над различными областями мозга, мы имеем возможность выявить локальные нарушения работы мозга, указывающие на поражение его ткани в определенном месте. Имея данные об изменении электроэнцефалограммы в процессе роста и развития ребенка – можем сделать вывод о темпах созревания мозга у данного конкретного маленького пациента.
Задачи для самостоятельной работы:
1. толшина коры головного мозга 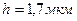 , удельное сопротивление коры
, удельное сопротивление коры 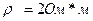 , коэффициент средней плотности пирамидных нейронов в коре
, коэффициент средней плотности пирамидных нейронов в коре 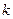
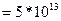 , среднее стандартное отклонение дипольного момента нейронов
, среднее стандартное отклонение дипольного момента нейронов 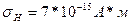 . Вычислите стандартное отклонение ЭЭГ пирмидных нейронов, при
. Вычислите стандартное отклонение ЭЭГ пирмидных нейронов, при 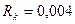 .
.
2. Найдите потенциал поля, созданного диполем в точке А, удаленной на расстояние 
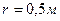 в направлении под углом
в направлении под углом 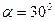 относительно электрического момента
относительно электрического момента 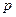 диполя. Среда – вода. Диполь образован зарядами
диполя. Среда – вода. Диполь образован зарядами 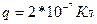 , расположенными на расстоянии
, расположенными на расстоянии 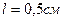 .
.
3. Диполь, образованный зарядами 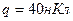 , плечом
, плечом 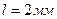 , свободно установилься в электрическом поле напряженностью
, свободно установилься в электрическом поле напряженностью 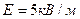 . Какую работу
. Какую работу 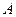 необходимо совершить, чтобы развернуть диполь на угол 300?
необходимо совершить, чтобы развернуть диполь на угол 300?
Контрольные тестовые задание на тему:
Литература:
- А.Н.Ремизов, А.Г.Максина, А.Я.Потапенко «Медицинская и биологическая физика», М., 2004 г..
- В.А.Тиманюк, Е.Н.Животова. Биофизика. М., 2004 г.
3. М.Е.Блохина, И.А. Эссаулова, Г.В.Мансурова. Руководство для лабораторных работ по медицинской и биологической физике. М. Дрофа.2002. – с. 242-246.

