




Наследственный дефект рецептора ЛПНП (к апо В/Е) (или белка апоВ-100) приводит к развитию распространённого наследственного заболевания — семейной гиперхолестеролемии.
При дефекте рецептора ЛПНП наблюдается триглицеролемия, при дефекте белка апоВ-100 – нет.
У гетерозигот (1:400) количество рецепторов ЛПНП на поверхности клеток снижено вдвое, а концентрация ХС в плазме примерно вдвое выше нормы (9-12 ммоль/л). ЛПНП фагоцитируются макрофагами. Нагруженные избытком ХС и других липидов, макрофаги откладываются в коже, сухожилиях и образуют ксантомы. Гиперхолестеролемия приводит к выраженному атеросклерозу, развитию ИБС и ранней смерти в результате инфаркта миокарда или инсульта.
У гомозигот (1:1000000) нет рецепторов к ЛПНП, концентрация ХС и ЛПНП в крови уже в раннем детском возрасте превышает норму в 5-6 раз (20-40 ммоль/л). Для пациентов характерно наличие не только ксантоматоза сухожилий, но и эруптивных ксантом на ягодицах, коленях, локтях, слизистой оболочке полости рта.
Избыток в крови ХС и ЛПНП способствует быстрому развитию атеросклероза и ИБС. Такие дети без экстренных мер лечения погибают в возрасте 5—6 лет.
ОБМЕН ЛПВП
ЛПВП выполняют 2 основные функции: они поставляют апо другим ЛП в крови и участвуют в так называемом «обратном транспорте ХС». ЛПВП синтезируются в печени и в небольшом количестве в тонком кишечнике в виде насцентных ЛПВП. Они имеют дисковидную форму, небольшой размер и содержат высокий процент белков и фосфолипидов. В печени в ЛПВП включаются апопротеины А, Е, С-II, ЛХАТ. В крови апо С-II и апо Е переносятся с ЛПВП на ХМ и ЛПОНП. насцентные ЛПВП практически не содержат ХС и ТГ и в крови обогащаются ХС, получая его из других ЛП и мембран клеток.
Для переноса ХС в ЛПВП существует сложный механизм. На поверхности ЛПВП находится фермент ЛХАТ — лецитин: холестерол-ацилтрансфераза. Этот фермент превращает ХС в ЭХС. Реакция активируется апо A-I, входящим в состав ЛПВП.
ЭХС перемещается внутрь ЛПВП. Таким образом, ЛПВП обогащаются ЭХС. ЛПВП увеличиваются в размерах, из дисковидных небольших частиц превращаются в частицы сферической формы, которые называют ЛПВП3, или «зрелые ЛПВП». ЛПВП3 частично обменивают ЭХС на ТГ, содержащиеся в ЛПОНП, ЛППП и ХМ. В этом переносе участвует «белок, переносящий эфиры холестерина» - апо D. Таким образом, часть ЭХС переносится на ЛПОНП, ЛППП, а ЛПВП3 за счёт накопления ТГ увеличиваются в размерах и превращаются в ЛПВП2.
Часть ЛПВП захватывается клетками печени, взаимодействуя со специфическими для ЛПВП рецепторами к апо А-1. На поверхности клеток печени ФЛ и ТГ ЛППП, ЛПВП2 гидролизуются печёночной липазой, что дестабилизирует структуру поверхности ЛП и способствует диффузии ХС в гепатоциты. ЛПВП2 в результате этого опять превращаются в ЛПВП3 и возвращаются в кровоток.
НАРУШЕНИЯ ЛПВП
Болезнь Тэнжи
Болеют аборигены острова Тэнжи. Наследственный дефект апо А, не синтезируются ЛПВП. Нарушается транспорт излишков ХС из тканей в печень. В крови низкий уровень ХС, ФЛ, много ТГ. Макрофаги фагоцитируют в тканях излишки ХС с образованием ксантом. Накопление ХС в печени, селезенке и других лимфоидных органах вызывает гепатоспленомегалию и лимфаденопатию. Может развиваться катаракта, полинейропатия и ренит. Миндалины из-за отложений ХС окрашены в оранжево-желтый цвет.
Список литературы
Берсенёв Алексей Вячеславович. Кандидатская диссертация: Трансплантация клеток эмбриональной печени и стволовых клеток костного мозга для коррекции дислипидемии и ранних стадий атерогенеза. М.: 2003.

ЛЕКЦИЯ № 13
Тема: Белая и бурая жировая ткань. Липолиз и липогенез.
Жировая ткань - белая и бурая: особенности химического состава, обмена белков, жиров, углеводов, энергетического метаболизма.
Жировая ткань – это разновидность соединительной ткани, составляет 15-20% веса мужчин и 20-25% веса женщин. Как и любая ткань, она состоит из клеток (адипоциты, макрофаги, тучные клетки и т.д.) и межклеточного вещества. Межклеточное вещество в жировой ткани представлено небольшим количеством волокон (коллагена и эластина) и основного вещества (гликозаминогликаны, протеоликаны).
Жировая ткань делиться на белую и бурую:
| Белая жировая ткань | Бурая жировая ткань | |
| Количество | Много | Мало у взрослого человека, много у новорожденного |
| Локализация | Есть везде. Основные жировые отложения имеются под кожей (в подкожножировой клетчатке), и вокруг внутренних органов, преимущественно в области живота (висцеральный жир). Толщина жирового слоя на животе может достигать 15-20 см (откладывается под кожей и в сальнике). Значительные жировые отложения имеются в забрюшинном пространстве, где располагаются почки, поджелудочная железа, аорта и другие органы. | В чистом виде: около почек и щитовидной железы. Смешанная жировая ткань: между лопатками, на грудной клетке и плечах. |
| Кровоснабжение | Слабое | Очень хорошее |
| Цвет | Бело-желтоватый | Бурый |
| Строение адипоцитов | В цитоплазме имеется одна большая капля жира, ядро сплющено, вместе с органеллами оттеснено к периферии | В цитоплазме много мелких капелек жира, ядро и органеллы расположены в центре клетки, имеется много митохондрий и цитохромов. |
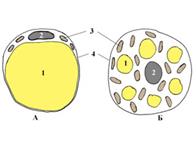
| 1. Жировой пузырек 2. Ядро клетки 3. Митохондрии 4. Оболочка клетки | |
| Функция | 1.Синтез липидов из углеводов 2.запасающая (95% ТГ в жировой ткани, 5% - во внутренних органах и костях); 3. теплоизоляционная; 4. механическая защита органов и тканей; 5. эндокринная (секреция: лептина, эстрогенов, ангиотензиногена, адипонектина, резистина, интерлейкина-6, ФНО-α); 6.Антибактериальная защита (комплементарные факторы: адипсин, комплемент С3, фактор В) 7.Регуляция воспаления (α1-кислый гликопротеин, гаптоглобин) 8.Накопление витаминов А, Д, Е 9.пассивное обезвреживание (депонирование) токсичных веществ 10. Участие в водном обмене Обеспечивает поддержание гомеостаза. | термогенез |
Развитие жировой ткани

| Жировая ткань развивается из мезенхимы с 30 недели эмбрионального развития. Мезенхимальная клетка превращается в липобласт, который в свою очередь, превращается в зрелую жировую клетку — адипоцит. Существует два периода активного увеличения количества адипоцитов: (1) период эмбрионального развития и (2) период полового созревания. В другие периоды жизни человека обычно размножения клеток-предшественников не происходит. Накопление жира идет только путем увеличения размеров уже существующих жировых клеток. Если количество жира в клетке достигает критической массы, клетки-предшественники получают сигнал, и начинают размножаться, давая рост новым жировым клеткам. У худого взрослого человека имеется около 35 миллиардов жировых клеток, у человека с выраженным ожирением до 125 миллиардов, то есть в 4 раза больше. Вновь образованные жировые клетки обратному развитию не подлежат, и сохраняются на всю жизнь. Если человек худеет, то они лишь уменьшаются в размерах. |
ХИМИЧЕСКИЙ СОСТАВ БЕЛОЙ ЖИРОВОЙ ТКАНИ
Жировая ткань содержит 65-85% ТГ, 22% воды, 5,8% белка, 15 ммоль/кг калия. Из жирных кислот 42—51% приходится на олеиновую, 22—31% - на пальмитиновую, 5—14% - на пальмитоолеиновую, 3—5 % - на миристиновую, 1—5 % - на линолевую кислоты.
Состав жировой ткани зависит от области тела, глубины слоя; он может также несколько отличаться у отдельных индивидуумов. Особенно подвергается изменениям содержание воды и белка. Чем глубже под поверхностью кожи жир расположен, тем больше он содержит насыщенных кислот. У новорожденных насыщенные жиры во всех слоях содержатся в одинаковом количестве.
ОСОБЕННОСТИ МЕТАБОЛИЗМА БЕЛОЙ ЖИРОВОЙ ТКАНИ
Энергетический обмен низкий, преимущественно анаэробный, ткань потребляет мало кислорода. Энергия АТФ в основном тратится на транспорт жирных кислот через клеточные мембраны (с участием карнитина).
Белковый обмен низкий, белки синтезируются адипоцитами преимущественно для собственных нужд. На экспорт в жировой ткани синтезируются лептин, белки острой фазы воспаления (α1-кислый гликопротеин, гаптоглобин), компоненты системы комплимента (адипсин, комплемент С3, фактор В), интерлейкины.
Углеводный обмен. Невысокий, преобладает катаболизм. Углеводный обмен в жировой ткани тесно связан с липидным.
Липидный обмен
Жировая ткань стоит на 2 месте по обмену липидов после печени. Здесь происходят реакции липолиза и липогенеза.
Липогенез. В жировой ткани синтез липидов идет в абсорбтивный период по глицерофосфатному пути. Процесс стимулируется инсулином.
Этапы липогенеза:
1. Под действием инсулина на рибосомах стимулируется синтез ЛПЛ.
2. ЛПЛ выходит из адипоцита и фиксируется на поверхности стенки капилляра с помощью гепарансульфата.
3. ЛПЛ гидролизует ТГ в составе липопротеинов

4. Образовавшийся глицерин уноситься кровью в печень.
5. Жирные кислоты из крови транспортируются в адипоцит.
6. Кроме поступающих из вне экзогенных жирных кислот, в адипоците синтезируются жирные кислоты из глюкозы. Процесс стимулируется инсулином.
7. Жирные кислоты в адипоците под действием Ацил-КоА синтетазы превращаются в Ацил-КоА.

7. Глюкоза поступает в адипоцит с участием ГЛЮТ-4 (активатор инсулин).
8. В адипоците глюкоза вступает в гликолиз с образованием ФДА (активатор инсулин).
9. В цитоплазме ФДА восстанавливается глицерол-ф ДГ до глицерофосфата:

Так как в жировой ткани нет глицерокиназы, глицерофосфат образуется только из глюкозы (не может из глицерина).
10. В митохондриях глицерофосфат под действием глицеролфосфат ацилтрансферазы превращается в лизофосфатид:

11. В митохондриях лизофосфатид под действием лизофосфатид ацилтрансферазы превращается в фосфатид:

11. Фосфатид под действием фосфотидат фосфогидролазы превращается в 1,2-ДГ:
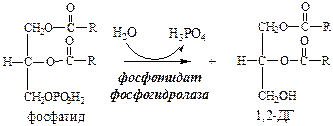
12. 1,2-ДГ под действием ацилтрансферазы превращается в ТГ:

13. Молекулы ТГ объединяются в крупные жировые капли.
 2. Липолиз. Липолиз в жировой ткани активируется при дефиците глюкозы в крови (постабсорбционный период, голодание, физическая нагрузка). Процесс стимулируется глюкагоном, адреналином, в меньшей степени СТГ и глюкокортикоидами.
2. Липолиз. Липолиз в жировой ткани активируется при дефиците глюкозы в крови (постабсорбционный период, голодание, физическая нагрузка). Процесс стимулируется глюкагоном, адреналином, в меньшей степени СТГ и глюкокортикоидами.
В результате липолиза концентрация свободных жирных кислот в крови возрастает в 2 раза.


ОСОБЕННОСТИ МЕТАБОЛИЗМА БУРОЙ ЖИРОВОЙ ТКАНИ
Энергетический обмен. Ткань потребляет много кислорода, активно окисляет глюкозу и жирные кислоты. Энергетический обмен высокий. При этом, АТФ образуется только в реакциях субстратного фосфорилирования (2 реакции гликолиза, 1 реакции ЦТК). Причина - разобщение в митохондриях белком термогенином (РБ-1) процессов окисления и фосфорилирования, низкая активность АТФ синтетазы, отсутствие дыхательного контроля со стороны АДФ. В бурой жировой ткани вся образующаяся при окислении энергия рассеивается в виде тепла (термогенез).
Термогенез в бурой жировая ткань активируется при переохлаждении СНС, а также при излишке липидов в крови, под действием лептина. Благодаря этому повышается температура тела и снижается концентрация липидов в крови. Отсутствие бурой жировой ткани у взрослых людей является причиной 10% всех случаев ожирения.
ЛЕКЦИЯ № 14
Тема: Обмен жирных кислот, регуляция
Строение жирных кислот
Жирными кислотами (ЖК) - называются карбоновые кислоты, которые образуются при гидролизе омыляемых липидов.
В основном к жирным кислотам относятся высшие карбоновые кислоты (содержащие 12 и более атомов С). Такие ЖК водонерастворимыми, они транспортируются в крови с помощью альбуминов, а в клетках - с помощью Z-белков.
ЖК человека и животных имеют некоторые особенности строения: 1) они монокарбоновые; 2) содержат четное количество атомов С, самая распространенная длина от 16 до 18 атомов С; 3) углеродный скелет неразветвлен; 4) бывают насыщенными и ненасыщенными (мононенасыщенными и полиненасыщенными); 5). двойные связи несопряжены (разделены метиленовыми мостиками) и имеют цис-конформацию.
| № | Жирная кислота | Индекс ЖК | ∆ ЖК | ω ЖК |
| Лауриновая | 12:0 | |||
| Миристиновая | 14:0 | |||
| Пальмитиновая | 16:0 | |||
| Пальмитолеиновая | 16:1 | ∆9 | ω9 | |
| Стеариновая | 18:0 | |||
| Олеиновая | 18:1 | ∆9 | ω9 | |
| Линолевая | 18:2 | ∆9,12 | ω6 | |
| Линоленовая | 18:3 | ∆9,12,15 | ω3 | |
| Октадекатетраеновая | 18:4 | ∆5,8,11,14 | ω3 | |
| Арахиновая | 20:0 | |||
| Гадолеиновая | 20:1 | ∆9 | ω9 | |
| Эйкозатриеновая | 20:3 | ∆8,11,14 | ω6 | |
| Арахидоновая | 20:4 | ∆5,8,11,14 | ω6 | |
| Эйкозапентаеновая | 20:5 | ∆5,8,11,14,17 | ω3 | |
| Бегеновая | 22:0 | |||
| Эруковая | 22:1 | ∆13 | ω9 | |
| Андреновая | 22:4 | ∆9,12,15,18 | ω6 | |
| Докозапентаеновая | 22:5 | ∆4,7,10,13,16 | ω6 | |
| Докозагексаеновая | 22:6 | ∆4,7,10,13,16,19 | ω3 | |
| Лигноцериновая | 24:0 | |||
| Невроновая | 24:1 | ∆15 | ω9 | |
| Цереброновая | 24:0 | α-гидрокси ЖК |
∆ ЖК – номера атомов С, у которых расположены двойные связи.
ω ЖК - число атомов С от последней двойной связи до конца цепи.
Биологическое значение ЖК
- полиеновые ЖК (арахидоновая, эйкозапентаеновая, эйкозатриеновая) используются для синтеза БАВ – эйкозаноидов (простагландинов, простациклинов, тромбоксанов, лейкотриенов, липоксинов).
- ЖК окисляются в аэробных условиях с образованием АТФ;
- ЖК являются структурным компонентом омыляемых липидов: восков, глицеролипидов, сфинголипидов, эфиров холестерина;
КАТАБОЛИЗМ ЖИРНЫХ КИСЛОТ
В живых организмах катаболизм ЖК протекает как в ферментативных так и в неферментативных реакциях.
· Ферментативный катаболизм ЖК происходит в основном в реакциях β-окисления. К побочным путям относиться ферментативное α- и ω-окисление ЖК, а также деградация ЖК в пероксисомах. Хотя эти побочные пути количественно менее важны, их нарушение может приводить к тяжелым заболеваниям.
· Неферментативный катаболизм ЖК протекает в реакциях перекисного окисления липидов (ПОЛ).
β-окисление ЖК
β-окисление — специфический путь катаболизма ЖК с неразветвленной средней и короткой углеводородной цепью. β-окисление протекает в матриксе митохондрий, при котором от С конца ЖК последовательно отделяется по 2 атома С в виде Ацетил-КоА. β-окисление ЖК происходит только в аэробных условиях и является источником большого количества энергии.
β-окисление ЖК активно протекает в красных скелетных мышцах, сердечной мышце, почках и печени. ЖК не служат источником энергии для нервных тканей, так как ЖК не проходят через гематоэнцефалический барьер, как и другие гидрофобные вещества.
β-окисление ЖК увеличивается в постабсорбтивный период, при голодании и физической работе. При этом концентрация ЖК в крови увеличивается в результате мобилизации ЖК из жировых ткани.
Активация ЖК
Активация ЖК происходит в результате образования макроэргической связи между ЖК и HSКоА с образованием Ацил-КоА. Реакцию катализирует фермент Ацил-КоА синтетаза:
RCOOH + HSKoA + АТФ → RCO~SКоА + АМФ+ PPн
Пирофосфат гидролизуется ферментом пирофосфатазой: Н4Р2О7 + Н2О → 2Н3РО4
Ацил-КоА синтетазы находятся как в цитозоле (на внешней мембране митохондрий), так и в матриксе митохондрий. Эти ферменты отличаются по специфичности к ЖК с различной длиной углеводородной цепи.
Транспорт ЖК
Транспорт ЖК в матрикс митохондрий зависит от длины углеродной цепи.
ЖК с короткой и средней длиной цепи (от 4 до 12 атомов С) могут проникать в матрикс митохондрий путём диффузии. Активация этих ЖК происходит ацил-КоА синтетазами в матриксе митохондрий.
ЖК с длинной цепью, сначала активируются в цитозоле (ацил-КоА синтетазами на внешней мембране митохондрий), а затем переносятся в матрикс митохондрий специальной транспортной системой с помощью карнитина. Карнитин поступает с пищей или синтезируется из лизина и метионина с участием витамина С.
· В наружной мембране митохондрий фермент карнитинацилтрансфераза I (карнитин-пальмитоилтрансфераза I) катализирует перенос ацила с КоА на карнитин с образованием ацилкарнитина;
· Ацилкарнитин проходит через межмембранное пространство к наружной стороне внутренней мембраны и транспортируется с помощью карнитинацилкарнитинтранслоказы на внутреннюю поверхность внутренней мембраны митохондрий;
· Фермент карнитинацилтрансфераза II катализирует перенос ацила с карнитина на внутримитохондриальный HSКоА с образованием Ацил-КоА;
· Свободный карнитин возвращается на цитозольную сторону внутренней мембраны митохондрий той же транслоказой.
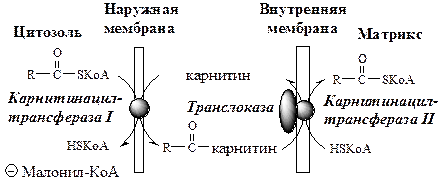
Реакции β-окисление ЖК
1. β-окисление начинается с дегидрирования ацил-КоА ФАД-зависимой Ацил-КоА дегидрогеназой с образованием двойной связи (транс) между α- и β-атомами С в Еноил-КоА. Восстановленный ФАДН2 окисляясь в ЦПЭ, обеспечивает синтез 2 молекул АТФ;
2. Еноил-КоА гидратаза присоединяет воду к двойной связи Еноил-КоА с образованием β-оксиацил-КоА;
3. β-оксиацил-КоА окисляется НАД зависимой дегидрогеназой до β-кетоацил-КоА. Восстановленный НАДН2, окисляясь в ЦПЭ, обеспечивает синтез 3 молекул АТФ;
4. Тиолаза с участием HКоА отщепляет от β-кетоацил-КоА Ацетил-КоА. В результате 4 реакций образуется Ацил-КоА, который короче предыдущего Ацил-КоА на 2 углерода. Образованный Ацетил-КоА окисляясь в ЦТК, обеспечивает синтез в ЦПЭ 12 молекул АТФ.
Затем Ацил-КоА снова вступает в реакции β-окисления. Циклы продолжаются до тех пор, пока Ацил-КоА не превратится в Ацетил-КоА с 2 атома С (если ЖК имела четное количество атомов С) или Бутирил-КоА с 3 атомами С (если ЖК имела нечетное количество атомов С).

