
| 
|

Поиск: Рекомендуем:  Почему я выбрал профессую экономиста
Почему я выбрал профессую экономиста
 Почему одни успешнее, чем другие
Почему одни успешнее, чем другие
 Периферийные устройства ЭВМ Периферийные устройства ЭВМ
 Нейроглия (или проще глия, глиальные клетки) Нейроглия (или проще глия, глиальные клетки)
Категории:
|
Многие коферменты и родственные им соединения являются производными аденозинмонофосфата
| ||||
| 2. Представление о биосинтезе пуриновых нуклеотидов. Роль ФРДФ. Происхождение атомов пуринового кольца. ИМФ как предшественник АМФ и ГМФ. Регуляция синтеза пуриновых нуклеотидов | |||||
Биосинтез пуриновых нуклеотидов
У человека и других млекопитающих пуриновые нуклеотиды синтезируются для обеспечения потребностей организма в мономерных предшественниках нуклеиновых кислот, а также в соединениях, выполняющих другие функции. У некоторых позвоночных (птицы, земноводные, рептилии) синтез пуриновых нуклеотидов несет дополнительную функцию – является частью механизма, с помощью которого выводятся излишки азота в виде мочевой кислоты; такие организмы называют урикотелическими. Организмы, у которых конечным продуктом азотистого обмена является мочевина (как у человека), называют уреотелическими. Поскольку урикотелические организмы удаляют «излишки» азота в виде мочевой кислоты, синтез пуриновых нуклеотидов у них идет более интенсивно, чем у уреотелических. В то же время пути синтеза пуриновых нуклеотидов de novo – общие для обеих групп организмов.
Пищевые пурины и пиримидины не являются незаменимыми пищевыми факторами и очень мало используются для синтеза нуклеиновых кислот тканей.
Пиримидиновые основания, не успевшие поступить в энтероциты, под действием микрофлоры кишечника расщепляются до NH3, CO2, β-аланина и β-аминоизобутирата.
В различных клетках организма синтезируется до 90% пуриновых и пиримидиновых нуклеотидов из простых предшественников de novo.
Фосфорибозилдифосфат (ФРДФ), или фосфорибозилпирофосфат (ФРПФ) занимает центральное место в синтезе как пуриновых, так и пиримидиновых нуклеотидов.
Образование 5-фосфорибозил-1-дифосфата (ФРДФ)
 Источником рибозо-5-фосфата могут быть: 1) пентозофосфатный путь превращения глюкозы; 2) катаболизм нуклеозидов, в ходе которого под действием нуклеозидфосфорилазы первоначально образуется рибозо-1-фосфат, а затем с помощью соответствующей мутазы фосфорный остаток переносится в 5-положение.
Происхождение атомов азота и углерода пуринового кольца
Источником рибозо-5-фосфата могут быть: 1) пентозофосфатный путь превращения глюкозы; 2) катаболизм нуклеозидов, в ходе которого под действием нуклеозидфосфорилазы первоначально образуется рибозо-1-фосфат, а затем с помощью соответствующей мутазы фосфорный остаток переносится в 5-положение.
Происхождение атомов азота и углерода пуринового кольца
 Ниже представлена схема реакций синтеза пуриновых нуклеотидов de novo.
Ниже представлена схема реакций синтеза пуриновых нуклеотидов de novo.
|
Переваривание нуклеиновых кислот пищи

| ||||

| |||||
Синтез АМФ и ГМФ из ИМФ
 1 – аденилосукцинатсинтаза; 2 – аденилосукциназа; 3 – ИМФ-дегидрогеназа; 4 – ГМФ-синтетаза.
Печень – основное место образования пуриновых нуклеотидов, откуда они могут поступать в ткани, не способные к их синтезу: эритроциты, полиморфноядерные лейкоциты (ПЯЛ) и частично мозг.
Регуляция синтеза пуриновых нуклеотидов
1 – аденилосукцинатсинтаза; 2 – аденилосукциназа; 3 – ИМФ-дегидрогеназа; 4 – ГМФ-синтетаза.
Печень – основное место образования пуриновых нуклеотидов, откуда они могут поступать в ткани, не способные к их синтезу: эритроциты, полиморфноядерные лейкоциты (ПЯЛ) и частично мозг.
Регуляция синтеза пуриновых нуклеотидов
 1 – ФРДФ-синтетаза; 2 – амидофосфорибозилтрансфераза; 3 – ИМФ-дегидрогеназа; 4 – аденилосукцинатсинтетаза.
1 – ФРДФ-синтетаза; 2 – амидофосфорибозилтрансфераза; 3 – ИМФ-дегидрогеназа; 4 – аденилосукцинатсинтетаза.
| |||||
| 3. Катаболизм пуриновых нуклеотидов. Нарушения метаболизма пуринов: подагра, синдром Леша-Найхана | |||||
Конечный продукт катаболизма пуринов у человека – мочевая кислота. При обследовании больных с наследственной формой недостаточности ферментных систем катаболизма пуринов установлено, что 99% мочевой кислоты образуется из субстратов нуклеозидфосфорилазы, функционирующей в цикле реутилизации пуринов. Пуриновые продукты нуклеозидфосфорилазной реакции – гипоксантин и гуанин – превращаются в мочевую кислоту; промежуточным продуктом является ксантин, образующийся в реакциях, катализируемых гуаназой и ксантиноксидазой в печени, тонком кишечнике и почках:
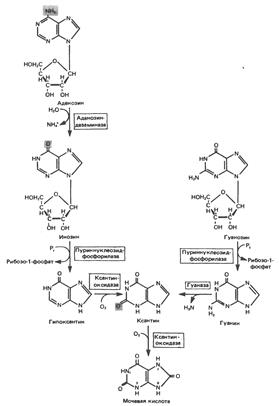 Ксантиноксидаза представляет собой важную мишень для фармакологического вмешательства при гиперурикемии и подагре.
У низших приматов и других млекопитающих (но не у человека) мочевая кислота гидролизуется уриказой до аллантоина – соединения, хорошо растворимого в воде:
Ксантиноксидаза представляет собой важную мишень для фармакологического вмешательства при гиперурикемии и подагре.
У низших приматов и других млекопитающих (но не у человека) мочевая кислота гидролизуется уриказой до аллантоина – соединения, хорошо растворимого в воде:
 У птиц и наземных рептилий уриказа отсутствует; в качестве конечных продуктов метаболизма азота (белков) и пуринов они экскретируют мочевую кислоту и гуанин. У этих организмов сформировалась урикотелическая система, позволяющая сохранить воду, ассоциированную с мочевой кислотой, при выделении последней в виде преципитата. Если бы конечным продуктом метаболизма азота у них была мочевина, сохранить гидратационную воду было бы невозможно, поскольку растворимость мочевины в воде достигает 10 моль/л (концентрация значительно выше той, которая может быть достигнута при концентрировании мочевины почками).
Метаболизм мочевой кислоты у человека (подагра)
Метаболизм мочевой кислоты у человека был изучен с применением изотопно-меченных мочевой кислоты, а также ее предшественников – глицина и формиата. По разведению инъецированного изотопа рассчитывали общее количество мочевой кислоты, находящейся в водной фазе организма. Этот параметр получил название «растворимый уратный пул».
Средняя величина данного показателя для здоровых взрослых мужчин составляла 1200 мг (разброс 866-1578 мг), а у здоровых женщин он колебался от 541 до 687 мг. У больных подагрой растворимый уратный пул был значительно выше и варьировал от 2000 до 4000 мг для пациентов без подагрических узлов, т.е. без отложений урата натрия в мягких тканях. При тяжелой форме подагры, сопровождающейся образованием узлов, растворимый уратный пул достигал величины 31000 мг. Скорость его обновления у здоровых людей составляет 600 мг за 24 ч.
18-20% удаляемой из организма мочевой кислоты распадается до СО2 и аммиака и выделяется через кишечник. Некоторое количество уратов экскретируется с желчью и подвергается деградации кишечной микрофлорой. Следует отметить, что распад мочевой кислоты до СО2 и NH3 у человека не связан с жизнедеятельностью кишечных бактерий.
Урат натрия легко фильтруется почечными клубочками млекопитающих, интенсивно реабсорбируется и частично экскретируется в проксимальных канальцах, затем секретируется в петле Генле и, вероятно, снова реабсорбируется в дистальных канальцах. За сутки здоровым человеком выделяется 400-600 мг мочевой кислоты.
Подагра – распространённое заболевание, в разных странах ею страдают от 0,3 до 1,7% населения. А поскольку сывороточный фонд уратов у мужчин в 2 раза больше, чем у женщин, то они и болеют чаще. Как правило, подагра генетически детерминирована и носит семейный характер. Она вызвана нарушениями в работе ФРДФ-синтетазы. К другим характерным проявлениям подагры относят нефропатию, при которой наблюдают образование уратных камней в мочевыводящих путях.
У птиц и наземных рептилий уриказа отсутствует; в качестве конечных продуктов метаболизма азота (белков) и пуринов они экскретируют мочевую кислоту и гуанин. У этих организмов сформировалась урикотелическая система, позволяющая сохранить воду, ассоциированную с мочевой кислотой, при выделении последней в виде преципитата. Если бы конечным продуктом метаболизма азота у них была мочевина, сохранить гидратационную воду было бы невозможно, поскольку растворимость мочевины в воде достигает 10 моль/л (концентрация значительно выше той, которая может быть достигнута при концентрировании мочевины почками).
Метаболизм мочевой кислоты у человека (подагра)
Метаболизм мочевой кислоты у человека был изучен с применением изотопно-меченных мочевой кислоты, а также ее предшественников – глицина и формиата. По разведению инъецированного изотопа рассчитывали общее количество мочевой кислоты, находящейся в водной фазе организма. Этот параметр получил название «растворимый уратный пул».
Средняя величина данного показателя для здоровых взрослых мужчин составляла 1200 мг (разброс 866-1578 мг), а у здоровых женщин он колебался от 541 до 687 мг. У больных подагрой растворимый уратный пул был значительно выше и варьировал от 2000 до 4000 мг для пациентов без подагрических узлов, т.е. без отложений урата натрия в мягких тканях. При тяжелой форме подагры, сопровождающейся образованием узлов, растворимый уратный пул достигал величины 31000 мг. Скорость его обновления у здоровых людей составляет 600 мг за 24 ч.
18-20% удаляемой из организма мочевой кислоты распадается до СО2 и аммиака и выделяется через кишечник. Некоторое количество уратов экскретируется с желчью и подвергается деградации кишечной микрофлорой. Следует отметить, что распад мочевой кислоты до СО2 и NH3 у человека не связан с жизнедеятельностью кишечных бактерий.
Урат натрия легко фильтруется почечными клубочками млекопитающих, интенсивно реабсорбируется и частично экскретируется в проксимальных канальцах, затем секретируется в петле Генле и, вероятно, снова реабсорбируется в дистальных канальцах. За сутки здоровым человеком выделяется 400-600 мг мочевой кислоты.
Подагра – распространённое заболевание, в разных странах ею страдают от 0,3 до 1,7% населения. А поскольку сывороточный фонд уратов у мужчин в 2 раза больше, чем у женщин, то они и болеют чаще. Как правило, подагра генетически детерминирована и носит семейный характер. Она вызвана нарушениями в работе ФРДФ-синтетазы. К другим характерным проявлениям подагры относят нефропатию, при которой наблюдают образование уратных камней в мочевыводящих путях.
| Синдром Леша-Найхана В некоторых случаях гиперэкскреция уратов (более 600 мг мочевой кислоты за 24 ч) может носить вторичный характер, т.е. быть следствием заболеваний, характеризующихся усилением обмена в тканях (например, рак или псориаз). У ряда пациентов идентифицированы дефекты ферментных систем, например ФРПФ-синтетазы (устойчивость к ингибированию конечными продуктами и повышенная активность), гипоксантингуанин-фосфорибозилтрансферазы (ГГФРТ) (уменьшение количества фермента или его полное отсутствие, синдром Леша-Найхана). К этой же категории можно отнести недостаточность глюкозо-6-фосфатазы (синдром Гирке). Имеется также группа больных с идиопатической гиперурикемией, которую следует рассматривать как гетерогенную до тех пор, пока не будет выяснена молекулярная основа метаболических дефектов. Синдром Леша-Найхана (полное отсутствие ГГФРТ) наследуется как сцепленный с X-хромосомой рецессивный признак. Болезнь характеризуется корковым параличом, сопровождающимся хореоатетозом, судорогами, стремлением к членовредительству и тяжелой гиперурикемией. Кроме того, наблюдается образование камней мочевой кислоты. Матери больных детей гетерозиготны и мозаичны в отношении ГГФРТазной недостаточности, у них часто обнаруживается гиперурикемия, но без неврологических симптомов. Частичная недостаточность ГГФРТазы, вызванная мутациями соответствующего гена, встречается и у мужчин. Для таких больных характерна тяжелая гиперурикемия, не сопровождающаяся существенными неврологическими нарушениями. Избыточное образование пуринов у пациентов с недостаточностью ГГФРТ связано с увеличенной внутриклеточной концентрацией ФРПФ, что по-видимому является результатом уменьшения потребления ФРПФ на пути регенерации пуриновых нуклеотидов. Биохимическая основа неврологических отклонений при синдроме Леша-Найхана неизвестна. | ||||
| 4 Синтез пиримидиновых нуклеотидов | |||||
Фонд пиримидиновых нуклеотидов, подобно пуриновым нуклеотидам, в основном синтезируется из простых предшественников de novo, и только 10-20% от общего количества образуется по «запасным» путям из азотистых оснований или нуклеозидов.
В отличие от синтеза пуринов, где формирование гетероциклического основания осуществляется на остатке рибозо-5-фосфата, пиримидиновое кольцо синтезируется из протсых предшественников: глутамина, СО2 и аспарагиновой кислоты и затем связывается с рибозо-5фосфатом, полученным от ФРДФ.
Процесс протекает в цитозоле клеток. Синтез ключевого пиримидинового нуклеотида – УМФ идёт с участием трёх ферментов, 2 из которых полифункциональны.
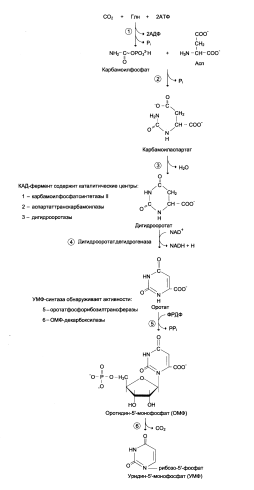
У млекопитающих ключевой, регуляторной реакцией в синтезе пиримидиновых нуклеотидов является синтез карбамоилфосфата из глутамина, СО2 и АТФ, в реакции, каталищзируемой карбамоилфосфатсинтетазой II (КФС II), которая протекает в цитозоле клеток:
 В реакции NH2-группа карбамоилфосфата образуется за счёт амидной группы глутамина, что отличает эту реакцию от реакции синтеза карбамоилфосфата в митохондриях в процессе синтеза мочевины из СО2, NH3 и АТФ с участием КФС I.
Карбамоилфосфат, использующийся на образование пиримидиновых нуклеотидов, является продуктом полифункционального фермента, который, наряду с активностью КФС II, содержит каталитические центры аспартаткарбамоилазы и дигидрооротазы. Этот фермент назвали КАД-фермент – по начальным буквам ферментативных активностей, которыми обладают отдельные каталитические домены этого белка.
Отщепляясь от КАД-фермента, дигидрооротат подвергается дегидрированию НАД-зависимой дигидрооротатдегидрогеназой и превращается в свободное пиримидоновое основание – оротовую кислоту, или оротат.
В цитозоле оротат становится субстратом бифункционального фермента – УМФ-синтетазы, которая обнаруживает оротатфосфорибозилтрансферазную и ОМФ-декарбоксилазную активности.
Первоначально фосфорибозильный остаток от ФРДФ переносится на оротат и образуется нуклеотид – оротидин-5´-монофосфат (ОМФ), декарбоксилирование которого даёт уридин-5-монофосфат (УМФ).
Биосинтез УМФ
В реакции NH2-группа карбамоилфосфата образуется за счёт амидной группы глутамина, что отличает эту реакцию от реакции синтеза карбамоилфосфата в митохондриях в процессе синтеза мочевины из СО2, NH3 и АТФ с участием КФС I.
Карбамоилфосфат, использующийся на образование пиримидиновых нуклеотидов, является продуктом полифункционального фермента, который, наряду с активностью КФС II, содержит каталитические центры аспартаткарбамоилазы и дигидрооротазы. Этот фермент назвали КАД-фермент – по начальным буквам ферментативных активностей, которыми обладают отдельные каталитические домены этого белка.
Отщепляясь от КАД-фермента, дигидрооротат подвергается дегидрированию НАД-зависимой дигидрооротатдегидрогеназой и превращается в свободное пиримидоновое основание – оротовую кислоту, или оротат.
В цитозоле оротат становится субстратом бифункционального фермента – УМФ-синтетазы, которая обнаруживает оротатфосфорибозилтрансферазную и ОМФ-декарбоксилазную активности.
Первоначально фосфорибозильный остаток от ФРДФ переносится на оротат и образуется нуклеотид – оротидин-5´-монофосфат (ОМФ), декарбоксилирование которого даёт уридин-5-монофосфат (УМФ).
Биосинтез УМФ
|
Синтез ЦТФ из УТФ
 Катаболизм пиримидиновых оснований
Катаболизм пиримидиновых оснований

| ||||
| 5. Регуляция синтеза пиримидинов. Конечные продукты распада пиримидинов. Нарушения метаболизма пиримидинов | |||||
Регуляторным ферментом в синтезе пиримидиновых нуклеотидов является полифункциональный КАД-фермент. УМФ и пуриновые нуклеотиды аллостерически ингибируют, а ФРДФ активирует его карбамоилсинтетазную активность, тогда как активность аспартаттранскарбамоилазного домена ингибирует ЦТФ, но активирует АТФ:
 Этот способ регуляции позволяет предотвратить избыточный синтез не только УМФ, но и всех других пиримидиновых нуклеотидов и обеспечить сбалансированное образование всех четырёх основных пуриновых и пиримидиновых нуклеотидов, необходимых для синтеза РНК.
Нарушения обмена пиримидиновых нуклеотидов
Описано несколько нарушений, связанных со снижением активности ферментов обмена пиримидиновых нуклеотидов. Одно из них – оротацидурия – вызвано дефектом в работе второго бифункционального фермента синтеза нуклеотидов de novo – УМФ-синтазы, два других обнаружены в процессе катаболизма пиримидинов.
Этот способ регуляции позволяет предотвратить избыточный синтез не только УМФ, но и всех других пиримидиновых нуклеотидов и обеспечить сбалансированное образование всех четырёх основных пуриновых и пиримидиновых нуклеотидов, необходимых для синтеза РНК.
Нарушения обмена пиримидиновых нуклеотидов
Описано несколько нарушений, связанных со снижением активности ферментов обмена пиримидиновых нуклеотидов. Одно из них – оротацидурия – вызвано дефектом в работе второго бифункционального фермента синтеза нуклеотидов de novo – УМФ-синтазы, два других обнаружены в процессе катаболизма пиримидинов.
| Клинически наиболее характерное следствие оротацидурии – мегалобластная анемия, вызванная неспособностью организма обеспечить нормальную скорость деления клеток эритроцитарного ряда. |
|
|
|
|
|
|
Дата добавления: 2016-11-23; Мы поможем в написании ваших работ!; просмотров: 1721 | Нарушение авторских прав
Лучшие изречения:
 | 4383 -
| 4383 - 