




Если нагреть раствор ДНК выше температуры 90°С или сдвинуть рН в резко щелочную или резко кислую стороны, то водородные связи между нитями ДНК разрушаются и двойная спираль расплетается. Происходит денатурация ДНК или, по-другому, плавление. Если удалить агрессивный фактор, то происходит ренатурация или отжиг. При отжиге нити ДНК "отыскивают" комплементарные участки друг у друга и, в конце концов, вновь сворачиваются в двойную спираль.
Если в одной "пробирке" провести плавление и отжиг смеси ДНК, например, человека и мыши, то некоторые участки цепей ДНК мыши будут воссоединяться с комплементарными участками цепей ДНК человека с образованием гибридов. Число таких участков зависит от степени родства видов. Чем ближе виды между собой, тем больше участков комплементарности нитей ДНК. Это явление называется гибридизация ДНК-ДНК.
Если в растворе присутствует РНК, то можно осуществить гибридизацию ДНК-РНК. Такая гибридизация помогает установить близость определенных последовательностей ДНК с какой-либо РНК.
Гибридизация ДНК-ДНК и ДНК-РНК используется как эффективное средство в молекулярной генетике, судебной медицине, антропологии для установления генетического родства между видами.
Репликация происходит перед делением клетки
Репликация (синтез) ДНК происходит не беспорядочно, а в строго определенный период жизни клетки. Всего выделяют 4 фазы клеточного цикла: митоз (М), синтетическую (S), пресинтетическую (G1, от англ. gap – интервал), постсинтетическую (G2).
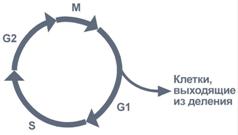
Фазы клеточного цикла
Важное участие в регуляции смены фаз клеточного цикла занимают циклины – белки массой 35-90 кДа, уровень которых меняется в ходе клеточного цикла. По функции циклины – это активаторные субъединицы ферментов циклин-зависимых киназ (ЦЗК). Активные комплексы циклин-ЦЗК фосфорилируют внутриклеточные белки, изменяя их активность. Этим обеспечивается продвижение по клеточному циклу.
Синтез (репликация, удвоение) ДНК происходит в S-фазу клеточного цикла, когда клетка готовится к делению. Механизм репликации, как установили Мэтью Мезельсон и Франклин Сталь в 1957 г, полуконсервативный, т.е. на каждой нити материнской ДНК синтезируется дочерняя копия.
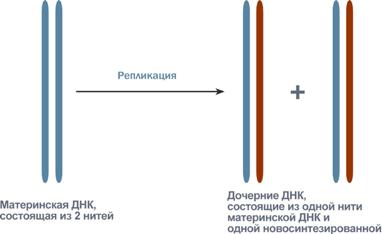
Схема полуконсервативного способа синтеза ДНК
Как любой матричный биосинтез, репликация требует наличия нескольких компонентов:
· матрица – в ее роли выступает материнская нить ДНК,
· растущая цепь – дочерняя нить ДНК,
· субстраты для синтеза – dАТФ, dГТФ, dЦТФ, ТТФ,
· источник энергии – dАТФ, dГТФ, dЦТФ, ТТФ,
· ферменты.
Синтез ДНК начинается в определенных участках, получивших название точка ori (англ. origin – начало). На каждой ДНК млекопитающих точек ori насчитывается до 100. Репликация распространяется от этих участков в обе стороны по нитям ДНК с образованием репликативных " пузырей ". В каждом таком "пузыре" имеются две репликативные "вилки", в которых происходит расплетание, раскручивание и непосредственный синтез ДНК. При этом репликативные вилки удаляются друг от друга. В целом репликация всех ДНК у эукариот заканчивается за 9 часов.
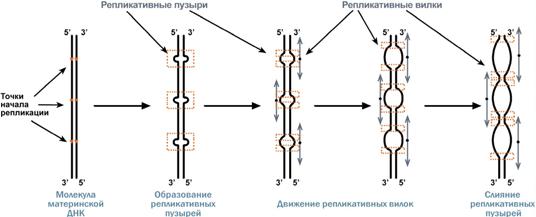
Схема возникновения репликативных пузырей и движения репликативных вилок
(нити дочерней ДНК не показаны). Стрелки - направление движения репликативных вилок.
Синтез новой цепи ДНК идет в направлении от 5'-конца к 3'-концу, т.е. 5'-конец новой ДНК остается свободным, каждый следующий нуклеотид своей 5'-гидроксильной группой присоединяется к 3'-гидроксильной группе предыдущего нуклеотида со скоростью порядка 100 штук в секунду.
В репликативной вилке в направлении 5'→3' непрерывно (т.е. обычным заурядным присоединением последующих нуклеотидов к предыдущим через С3 и С5) синтезируется только одна нить, а именно та, для которой направление синтеза совпадает с направлением движения репликативной вилки и соответствует направлению материнской нити 3'→5'. По мере расплетания ДНК и движения репликативной вилки на этой материнской нити открываются участки, где возможно безостановочное удлинение ведущей дочерней нити.
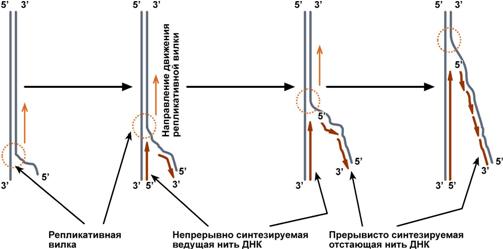
Схема движения репликативной вилки и синтеза ведущей и отстающей нитей ДНК
Направление 5'→3' для другой дочерней нити ДНК противоположно движению репликативной вилки. Поэтому синтез этой отстающей нити (в направлении 5'→3') возможен только после расплетания части ДНК и освобождения участка для синтеза.
Таким образом, синтез дочерней ДНК идет фрагментарно. По имени японского исследователя синтезируемые на отстающей цепи отрезки ДНК назвали фрагменты Оказаки.
Синтез ДНК не настолько сложен как кажется
В хронологическом порядке события репликации развертываются примерно следующим образом:
1. ДНК-топоизомеразы, находясь перед репликативной вилкой, разрезают молекулу ДНК для облегчения ее расплетания и раскручивания.
2. ДНК-хеликазы, следуя за топоизомеразами, раскручивают и расплетают молекулу ДНК.
3. ДНК-связывающие белки (ДСБ) связывают расплетенные нити ДНК и стабилизируют их, не допуская обратного "слипания" друг с другом.
4. ДНК-полимераза δ (греч.: δ – дельта), согласовано со скоростью движения репликативной вилки, осуществляет синтез ведущей цепи дочерней ДНК в направлении 5'→3' на матрице материнской нити ДНК по направлению от ее 3'-конца к 5'-концу (скорость до 100 пар нуклеотидов в секунду).
Этим события на данной материнской нити ДНК ограничиваются.
Далее описан синтез отстающей цепи дочерней ДНК.
5. Непосредственно сразу после расплетания и стабилизации другой нити материнской молекулы к ней присоединяется ДНК-полимераза α (α- альфа) и в направлении 5'→3' синтезирует праймер (РНК-затравку) – последовательность РНК на матрице ДНК длиной от 10 до 200 нуклеотидов. После этого фермент удаляется с нити ДНК.
Вместо ДНК-полимеразы α к 3'-концу праймера присоединяется ДНК-полимераза ε.
6. ДНК-полимераза ε (греч.: ε – эпсилон) как бы продолжает удлинять праймер, но в качестве субстрата встраивает дезокси рибонуклеотиды (в количестве 150-200 нуклеотидов). В результате образуется цельная нить из двух частей – РНК (т.е. праймер) и ДНК. ДНК-полимераза ε работает до тех пор, пока не встретит праймер предыдущего фрагмента Оказаки (синтезированный чуть ранее). После этого данный фермент удаляется с цепи.
7. ДНК-полимераза β (греч.: β – бета) встает вместо ДНК-полимеразы ε, движется в том же направлении (5'→3') и удаляет рибонуклеотиды праймера, одновременно встраивая дезокси рибонуклеотиды на их место. Фермент работает до полного удаления праймера, т.е. пока на его пути не встанет дезоксирибонуклеотид (еще более ранее синтезированный ДНК-полимеразой ε). Связать результат свой работы и впереди стоящую ДНК фермент не в состоянии, поэтому он сходит с цепи.
В результате на матрице материнской нити "лежит" фрагмент дочерней ДНК. Он называется фрагмент Оказаки.
8. ДНК-лигаза производит сшивку двух соседних фрагментов Оказаки, т.е. 5'-конца отрезка, синтезированного ДНК-полимеразой ε, и 3'-конца цепи, встроенного ДНК-полимеразой β.
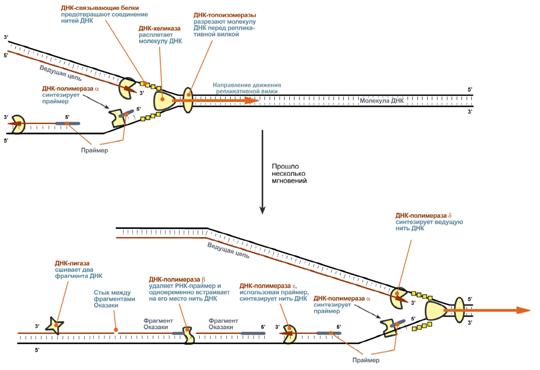
Схема репликативной вилки и функции ферментов репликации

