




Реферат
на тему:
«Можливості ДНК»
Виконала:
студентка I курсу ХТ-11-1
Побута Зоряна
Перевірив:
Доцент кафедри хімічної технології
Ганзюк А.Я..
План
1.Дизоксирибонуклеїнова кислота
2. Історія дослідження
3.Нуклеотиди
4.Подвійна спіраль
5.Утворення зв'язків між основами
6.Альтернативні форми подвійної спіралі
7.Суперскрученість
8.Структури на кінцях хромосом
9. Метилювання ДНК
10.Пошкодження ДНК
11. Біологічна роль ДНК
12. Використання ДНК в технології
13.Генна інженерія
14.Біоінформатика
15.Історія і антропологія
Висновок
1.Дезоксирибонуклеїнова кислота
Дезоксирибонуклеїнова кислота (ДНК) — один із двох типів природних нуклеїнових кислот, який забезпечує зберігання, передачу з покоління в покоління і реалізацію генетичної програми розвитку й функціонування живих організмів. Основна роль ДНК в клітинах — довготривале зберігання інформації про структуру РНК і білків.
У клітинах еукаріотів (наприклад, тварин, рослин або грибів) ДНК знаходиться в ядрі клітини в складі хромосом, а також в деяких клітинних органелах(мітохондріях і пластидах). У клітинах прокаріотів (бактерій і архей) кільцева або лінійна молекула ДНК, так званий нуклеоїд, знаходиться в цитоплазмі і прикріплена зсередини до клітинної мембрани. У них і у нижчих еукаріот (наприклад дріжджів) зустрічаються також невеликі автономні кільцеві молекули ДНК, так звані плазміди. Крім того, одно- або дволанцюжкові молекули ДНК можуть утворювати геном ДНК-вірусів.
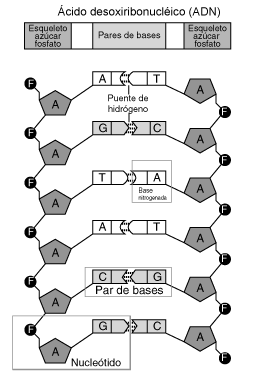
З хімічної точки зору, ДНК — це довга полімерна молекула, що складається з послідовності блоків — нуклеотидів. Кожний нуклеотид складається з азотистої основи, цукру (дезоксирибози) і фосфатної групи (або гомологічної арсеноїдної). Зв'язки між нуклеотидами в ланцюжку утворюються за рахунок дезоксирибози і фосфатної групи. У переважній більшості випадків (окрім деяких вірусів, що містять одноланцюжкові ДНК) макромолекула ДНК складається з двох ланцюжків, орієнтованих азотистими основами один проти одного. Ця дволанцюжкова молекула утворює спіраль. В цілому структура молекули ДНК отримала назву «подвійної спіралі».

У ДНК зустрічається чотири види азотистих основ (аденін, гуанін, тимін і цитозин) (виняток становлять випадки пізніших модифікацій нуклеотидів, наприклад метилювання). Азотисті основи одного з ланцюжків сполучені з азотистими основами іншого ланцюжка водневими зв'язками згідно з принципом комплементарності: аденін з'єднується тільки з тиміном, гуанін — тільки з цитозином. Послідовність нуклеотидів дозволяє «кодувати» інформацію про різні типи РНК, найважливішими з яких є інформаційні, або матричні (мРНК), рибосомальні (рРНК) і транспортні (тРНК). Всі ці типи РНК синтезуються на матриці ДНК (тобто за рахунок копіювання послідовності ДНК у послідовність макромолекули, що синтезується) у процесі транскрипції і беруть участь у біосинтезі білків (процесах сплайсингу і трансляції). Крім кодуючих послідовностей, ДНК клітини містить послідовності, що виконують регуляторні і структурні функції. Ділянки кодуючої послідовності разом із регуляторними ділянками називаються генами.
У геномах еукаріотів містяться також довгі послідовності без очевидної функції (некодуючі послідовності, інтрони). Також у складі геному досить поширені генетичні паразити — транспозони і вірусні або схожі на них послідовності.
Розшифровка структури ДНК (виконана в 1953 році) стала одним з поворотних моментів в історії біології. За видатний внесок у це відкриття Френсісу Кріку, Джеймсу Ватсону і Морісу Вілкінсу була присуджена Нобелівська премія з фізіології і медицини 1962 року.
2.Історія дослідження
ДНК була відкрита Іоганном Фрідріхом Мішером у 1869 році. Спочатку нова речовина отримала назву нуклеїн, а пізніше, коли Мішер визначив, що ця речовина володіє кислотними властивостями, речовина отримала назву нуклеїнова кислота. Біологічна функція нововідкритої речовини була неясна, і довгий час ДНК вважалася запасником фосфору в організмі. Більш того, навіть на початку 20 століття багато біологів вважали, що ДНК не має ніякого відношення до передачі інформації, оскільки будова молекули, на їхню думку, була дуже одноманітною і не могла містити закодовану інформацію.
Поступово було доведено, що саме ДНК, а не білки, як вважалося раніше, є носієм генетичної інформації. Одними з перших вирішальних доказів стали експерименти О. Евері, Коліна Мак-Леода і Маклін Мак-Карті (1944 рік) з трансформації бактерій. Їм вдалося показати, що за так звану трансформацію (придбання хвороботворних властивостей нешкідливою культурою у результаті додавання до неї мертвих хвороботворних бактерій) відповідає виділена з пневмококів ДНК. Експеримент американських учених Альфреда Хершу і Марти Чейз (1952 рік) з міченими радіоактивними ізотопами білками і ДНК бактеріофагів показали, що в заражену клітку передається тільки нуклеїнова кислота фага, а нове покоління фага містить такі ж білки і нуклеїнову кислоту, як і початковий фаг.
До 50-х років 20 століття точна будова ДНК, як і спосіб передачі спадкової інформації, залишалася невідомою. Хоч і було напевно відомо, що ДНК складається з кількох ланцюжків, що у свою чергу складаються з нуклеотидів, ніхто не знав точно, скільки цих ланцюжків і як вони сполучені.
3. Нуклеотиди
Структура подвійної спіралі ДНК була запропонована Френсісом Кріком і Джеймсом Ватсоном у 1953 році на основі рентгеноструктурних даних, отриманих Морісом Вілкінсом і Розаліндою Франклін, і «правил Чаргаффа», згідно з якими в кожній молекулі ДНК дотримуються строгі співвідношення, що зв'язують між собою кількість азотистих основ різних типі. Пізніше запропонована Ватсоном і Кріком модель будови ДНК була доведена, а їхня робота відмічена Нобелівською премією з фізіології і медицини 1962 року. Серед одержувачів не було Розалінди Франклін, що померла на той час, оскільки премія не присуджується посмертно.
У відомій доповіді 1957 року, Крік окреслив основи так званої «Центральної догми» молекулярної біології, яка передбачає взаємовідношення між ДНК, РНК і білками, та сформулював «адаптерну гіпотезу». Остаточне підтвердження механізму копіювання, запропонованого на основі спіральної структури, було отримане в 1958 році за допомогою експерименту Мезельсона-Сталя. Подальші роботи Кріка і його лабораторії показали, що генетичний код засновується на трійках основ, що не перекриваються, кодонах, що пізніше дозволило Гару Ґобінду Хорані, Роберту Голлі і Маршаллу Ніренбергу розшифрувати генетичний код. Ці відкриття позначають початок молекулярної біології.
Дезоксирибонуклеїнова кислота є біополімером (поліаніоном), мономерами якого є нуклеотиди. Кожен нуклеотид складається із залишку фосфорної кислоти, приєднаного за 5-положенням до цукру дезоксирибози, до якого також через глікозидний зв'язок (C—N) за 1-положенням приєднана одна з чотирьох азотистих основ. Саме наявність характерного цукру і складає одну з головних відмінностей між ДНК і РНК, зафіксовану в назвах цих нуклеїнових кислот (до складу РНК входить цукор рибоза). Приклад нуклеотиду — аденозин монофосфат — де основа, приєднана до фосфату і рибози, це аденін, показаний на малюнку.
Виходячи із структури молекул, основи, що входять до складу нуклеотидів, розділяють на дві групи: пуринові (аденін [A] і гуанін [Г]), утворені сполученими п'яти- і шестичленним гетероциклами і піримідинові (цитозин [Ц] і тимін [T]) — утворені одним шестичленним гетероциклом.
Як виняток, наприклад, у бактеріофага, в ДНК зустрічається п'ятий тип основ — урацил (У), піримідинова основа, що зазвичай входить до складу РНК замість тиміну і відрізняється від тиміну відсутністю метильної групи на кільці. Слід зазначити, що тимін і урацил не так строго приурочені до ДНК і РНК відповідно, як це вважалося раніше. Так, після синтезу деяких молекул РНК значне число урацилів у цих молекулах метилюєтся за допомогою спеціальних ферментів, перетворюючись на тимін. Це відбувається в транспортних і рибосомних РНК.


Тимін Аденін


Цитозин Гуанін
4. Подвійна спіраль
Полімер ДНК має досить складну структуру. Нуклеотиди ковалентно сполучені між собою в довгі полінуклеотидні ланцюжки. Ці ланцюжки в переважній більшості випадків (окрім деяких вірусів, що мають одноланцюжковий ДНК-геном), у свою чергу, попарно об'єднуються за допомогою водневих зв'язків у структуру, що отримала назву подвійної спіралі. Фосфатні групи формують фосфодіестерні зв'язки між третім і п'ятим атомами вуглецю сусідніх молекул дезоксирибози, в результаті взаємодії між 3-гідроксильною (3 —ОН) групою однієї молекули дезоксирибози і 5-фосфатною групою (5 —РО3) іншої. Асиметричні кінці ланцюжка ДНК називаються 3 (три-прайм) і 5 (п'ять-прайм). Полярність ланцюжка грає важливу роль при синтезі ДНК (подовження ланцюжка можливе тільки шляхом приєднання нових нуклеотидів до вільного 3 кінця).
Як вже було сказано вище, у переважної більшості живих організмів ДНК складається не з одного, а з двох полінуклеотидних ланцюжків. Ці два довгі ланцюжки закручені один навколо іншого у вигляді подвійної спіралі, що стабілізується водневими зв'язками, які утворюються між повернутими один до одного азотистими основами ланцюжків, що входять до неї. У природі ця спіраль, зазвичай, правозакручена. Напрями від 3 кінця до 5 кінця в двох ланцюжках, з яких складається молекула ДНК, протилежні (ланцюжки «антипаралельні» один одному). Ширина подвійної спіралі в її найпоширенішій B-формі складає від 22 до 24 А, або 2,2 — 2,4 нм, а довжина кожного нуклеотида 3,3 А (0,33 нм). Довжина всієї молекули залежить від виду організму, та може складати від десятків мікрон у деяких вірусів до кількох метрів (в одній хромосомі) у деяких рослин. Подібно до того, як в гвинтових сходах збоку можна побачити сходинки, на подвійній спіралі ДНК в проміжках між фосфатним остовом молекули можна бачити ребра основ, кільця яких розташовані в площині, перпендикулярній до подовжньої осі макромолекули. У подвійній спіралі розрізняють малу (12 А) і велику (22 А) борозенки. Білки, наприклад, фактори транскрипції, які приєднуються до певних послідовностей в дволанцюжковій ДНК, зазвичай взаємодіють з краями основ у великій борозенці, де вони доступніші.  .
.
5. Утворення зв’язків між основами
Кожна основа на одному з ланцюжків зв'язується з однією певною основою на іншому ланцюжку. Таке специфічне зв'язування називається комплементарним. Пуринові основи комплементарні піримідиновим (тобто, здатні до утворення водневих зв'язків з ними): аденін утворює зв'язки тільки з тиміном, а цитозин — з гуаніном. У подвійній спіралі ланцюжки також зв'язані за допомогою гідрофобної взаємодії і стекінгу, які не залежать від послідовності основ ДНК.
Комплементарність подвійної спіралі означає, що інформація, що міститься в одному ланцюжку, міститься і в іншому ланцюжку. Оборотність і специфічність взаємодій між комплементарними парами основ важлива для реплікації ДНК і решти всіх функцій ДНК в живих організмах.
Оскільки водневі зв'язки нековалентні, вони легко розриваються і відновлюються. Ланцюжки подвійної спіралі можуть розходитися як замок-змійка під дією ферментів (гелікази) або при високій температурі.
Різні пари основ утворюють різну кількість водневих зв'язків. A-T зв'язані двома, Г-Ц — трьома водневими зв'язками, тому на розрив Г-Ц потрібно більше енергії. Відсоток Г-Ц пар і довжина молекули ДНК визначають кількість енергії, необхідної для дисоціації ланцюжків: довгі молекули ДНК з великим Г-Ц більш «тугоплавкі».


6. Альтернатвні форми подвійної спіралі
ДНК може існувати в кількох можливих конформаціях. Зараз ідентифіковані та описані такі: A-ДНК, B-ДНК, C-ДНК, D-ДНК, E-ДНК, H-ДНК, L-ДНК, P-ДНК і Z-ДНК. Проте, тільки A-, B- і Z- форми ДНК спостерігалися в природних біологічних системах. Конформація, яку приймає ДНК, залежить від послідовності ДНК, величини та напрямку суперскрученості, хімічних модифікації основ і концентрації хімічних речовин у розчині, перш за все концентрацій іонів металів і поліамінів. Із всіх конформацій, B-форма, описана вище, є найзагальнішою формою за умов, що властиві більшості клітин. Альтернативні конформації подвійної спіралі відрізняються своєю геомертією та розмірами.
A-форма — ширша правостороння спіраль, з дрібнішою і ширшою малою борозенкою і вужчою і глибшою великою борозенкою. Ця форма зустрічається за нефізіологічними умовами в зневоднених зразках ДНК, крім того вона, ймовірно, зустрічається в живих клітинах у гібридних комплексах ланцюжків ДНК і РНК, та в комплексах ферментної ДНК. Сегменти ДНК із хімічно зміненими (метильованими) основами можуть проходити через більші конформаційні зміни і приймають Z-форму. Тут, ланцюжки закручаються в ліву подвійну спіраль, на відміну від правої спіралі B-форми. Ці структури можуть розпізнаватися специфічними Z-ДНК зв'язуючими білками і можуть бути залучені до регуляції транскрипції.

В залежності від концентрації іонів і нуклеотидного складу молекули, подвійна спіраль ДНК у живих організмах існує в різних формах. На малюнку (зліва направо) представлені A, B і Z форми
7. Супескручуваність
Суперскрученість ДНК — топологічна характеристика молекул ДНК. У «розслабленому» стані подвійної спіралі фрагменту ДНК, два ланцюжка, з яких він складається, обертаються навколо осі спіралі один раз на кожні 10,4 пар основ нуклеотидної послідовності ДНК. Проте, такий стан не є звичайним в живих клітинах, оскільки в процесі використання ДНК деякі ферменти додають або прибирають витки. Якщо фрагмент ДНК, кінці якого обернуті один навколо одного, замкнути у коло, а потім дозволити йому рухатися вільно, кільцева
ДНК отримала би нову форму, наприклад форму цифри вісім. Така зміна форми у порівнянні із круговою і є суперскрученістю.
Форма цифри вісім є найпростішим прикладом суперскрученості, і є формою, яку кругова ДНК приймає при додаванні одного витка навколо осі спіралі. Дві частини цифри вісім можіть обертатися або за годинниковою стрілкою, або проти годинникової стрілки одна щодо другої, залежно від того, чи спіраль закручена праворуч (надлишково закручена) або ліворуч (недозакручена). При додаванні кожного додаткового оберту навколо осі спіралі, частини молекули ДНК покажуть на один оберт більше.
Із малюнка можна побачити, що існують два типи суперскрученості, які, проте, топологічно рівнозначні: обертання ланцюжків в молекулі один навколо одного і перехрещення дволанцюжкових молекул. Формально суперскрученість є абстрактною математичною властивістю, що представляє собою суму обертів ланцюжків і перехрещень молекули:
S = T + W.
Додаткові оберти, в порівнянні із звичайним правим обертанням B-форми ДНК, приводять до позитивної суперскрученості, віднімання обертів — до негативної. Багато ферментів-топоізомераз додають або прибирають витки, чим змінюють топологію ДНК.
ДНК більшості організмів мають негативну суперскрученість. Хоча ДНК багатьох організмів лінійні, вони також негативно суперскручені, зокрема тому, що хромосоми найчастіше дуже великі і обертаються настільки повільно, що для ділянок в центрі здається, що кінці хромосом закріплені. В результаті, вони, можливо, не в змозі поширювати додаткові супервитки на всю хромосому, але центральна частина хромосом може бути так саме суперскручена, як і кругові молекули ДНК.
На кінцях лінійних хромосом є спеціалізовані структури ДНК, що називаються теломерами. Основна функція цих ділянок — підтримка цілісності кінців хромосом. Оскільки звичайна ДНК-полімераза не може реплікувати 3-кінці хромосом, спеціальний фермент —теломераза — після кожного поділу клітини додає до хромосом повторювані ділянки ДНК, теломери.
Теломери також захищають кінці ДНК від деградації екзонуклеазами і запобігають активації систем репарації, які запускаються у відповідь на розриви ДНК.
У клітинах людини теломери зазвичай представлені одноланцюжковою ДНК і складаються з декількох тисяч одиниць послідовності TTAГГГ, що повторюються. Ці послідовності з високим вмістом гуаніну стабілізують кінці хромосом, формуючи дуже незвичайні структури, які називають Г-квадруплексами і які складаються з чотирьох, а не двох взаємодіючих основ. Чотири гуанінових основи, всі атоми яких знаходяться в одній площині, утворюють пластинку, стабілізовану водневими зв'язками між основами і хелатованим у центрі іоном металу (найчастіше калію). Ці пластинки складаються стопкою одна над іншою.
8. Структури на кінцях хромосом
На кінцях хромосом можуть утворюватися і інші структури: основи можуть бути розташовані в одному ланцюжку або в різних паралельних ланцюжках. Окрім цих структур «стопок», теломери формують великі петлеподібні структури, звані Т-петлі або теломерні петлі. У них одноланцюжкова ДНК розташовується у вигляді широкого кільця, стабілізованого теломерними білками. У кінці Т-петлі одноланцюжкова теломерна ДНК приєднується до дволанцюжкової ДНК, порушуючи спаровування ланцюжків у цій молекулі й утворюючи зв'язки з одним із ланцюжків. Це триланцюжкове утворення називається D-петлею.
9. Метилюванння ДНК
За певних умов основи ДНК піддаються хімічним модифікаціям, які можуть бути успадковані без заміни послідовності ДНК, і, таким чином, є частиною епігенетичного коду. Найпоширенішим і найкраще описаним механізмом хімічних модифікацій є метилювання основ ДНК, цитозину у еукаріотів та цитозину і аденіну у бактерій.
Метилювання ДНК виявлене у всіх клітинах еукаріотів, проте середній рівень метилювання відрізняється у різних організмів, так унематоди метилювання цитозину майже не спостерігається, а у хребетних виявлений високий рівень метилювання — до 1 %. Відомо, що рівень метилювання цитозину впливає на експресію генів: ділянки гетерохроматину (що характеризуються відсутністю або низьким рівнем транскрипції) корелюють із рівнем метилювання. Наприклад, метилювання цитозину з утворенням 5-метилцитозину важливе для інактивації Х-хромосоми. Незважаючи на біологічну роль, 5-метилцитозин може спонтаннодезамінуватися, перетворюючись на тимін, тому метильований цитозин є джерелом підвищеного числа мутацій.
Крім контролю експресії генів та, в результаті, контролю клітинного циклу, бактерії використовують метилювання аденіну і цитозину для захисту проти патогенів у складі рестрикційно-модифікаційної системи.
Іншим добре описаним типом модифікацій основ є глікозування урацилу з утворенням «J-основи» в кінетопластидах.
10. Пошкодження ДНК
ДНК може пошкоджуватись різноманітними мутагенами, до яких належать окислюючі й алкілюючі речовини, а також високоенергетична електромагнітна радіація — ультрафіолетове й рентгенівське випромінювання. Тип пошкодження ДНК залежить від типу мутагена. Наприклад, ультрафіолет пошкоджує ДНК шляхом появи в ній димерів тиміну, які утворюються при формуванні ковалентних зв'язків між сусідніми основами.
Активні форми кисню, наприклад «вільні» радикали або перекис водню призводять до кількох типів пошкодження ДНК, включаючи модифікації основ, особливо гуанозину, а також дволанцюжкові розриви в ДНК. За деякими оцінками у кожній клітині людини близько 500 основ пошкоджуються окислюючими сполуками щодня. Серед різних типів пошкоджень найнебезпечніші — дволанцюжкові розриви, тому що вони важко репаруються і можуть призвести до втрат ділянок хромосом (делецій) і транслокацій.
Багато молекул мутагенів вставляються (інтеркалюються) між двома сусідніми парами основ. Більшість цих сполук, наприклад, бромистий етидій,дауноміцин, доксорубіцин і талідомід, мають ароматичну структуру. Для того, щоб ароматична сполука могла вміститися між основами, вони повинні розійтися, розплітаючи й порушуючи структуру подвійної спіралі. Ці зміни в структурі ДНК перешкоджують транскрипції і реплікації, викликаючи мутації. Тому інтеркалюючі речовини часто є канцерогенами, найвідоміші з яких — бензопірен, акридини, афлатоксини і бромистий етидій. Незважаючи на ці негативні властивості, в силу своєї здатності пригнічувати транскрипцію і реплікацію ДНК, деякі речовини, що інтеркалюють до ДНК, використовуються в хіміотерапії для пригнічення швидкого росту ракових клітин.
 (Інтеркальована хімічна сполука, що знаходиться в середині спіралі ДНК, — бензопірен, основний мутаген тютюнового диму.)
(Інтеркальована хімічна сполука, що знаходиться в середині спіралі ДНК, — бензопірен, основний мутаген тютюнового диму.)
11. Біологічна роль ДНК
ДНК є носієм генетичної інформації, записаної у вигляді нуклеотидної послідовності за допомогою генетичного коду. З молекулами ДНК зв'язані дві основоположні властивості живих організмів — спадковість і мінливість. У ході процесу, що називається реплікацією ДНК, утворюються дві копії початкового ланцюжка, які успадковуються дочірніми клітинами при поділі. Клітини, що утворилися таким чином, будуть генетично ідентичними. Генетична інформація, потрібна для життєдіяльності клітини, зчитується при експресії генів. У більшості випадків вона використовується для біосинтезу білків у процесах транскрипції (синтезу молекулРНК на матриці ДНК) і трансляції (синтезу білків на матриці РНК).
Послідовність нуклеотидів «кодує» інформацію про різні типи РНК: кодуючі — матричні або інформаційні (мРНК) — та некодуючі — рибосомальні (рРНК), транспортні (тРНК), каталітичні та інші. Всі ці типи РНК синтезуються на основі ДНК у процесі транскрипції. Їхня роль у біосинтезі білків та інших процесах життєдіяльності клітини різна. Матрична РНК містить інформацію про послідовність амінокислот у білку, рибосомальні РНК служать основою для рибосом (складних нуклеопротеїнових комплексів, основна функція яких — збірка білка з окремих амінокислот на основі мРНК), транспортні РНК доставляють амінокислоти до місця збірки білків — в активний центр рибосоми, що рухається по мРНК.
Генетичний код — набір правил розташування нуклеотидів в молекулах нуклеїнових кислот (ДНК і РНК), що надає всім живим організмам можливість кодування амінокислотної послідовності білків за допомогою послідовності нуклеотидів.
У ДНК використовується чотири нуклеотиди — аденін (А), гуанін (Г), цитозин (Ц) і тімін (T), які в україномовній літературі також часто позначаються буквами А, Г, Ц і Т відповідно. Ці букви складають «алфавіт» генетичного коду. У РНК використовуються ті ж нуклеотиди, за винятком тіміну, який замінений схожим нуклеотидом, — урацилом, який позначається буквою У (або У в україномовній літературі). У молекулах ДНК і РНК нуклеотиди складають ланцюжки і, таким чином, інформація закодована у вигляді послідовності генетичних «букв».
Для синтезу білків в природі використовуються 20 різних амінокислот. Кожен білок є ланцюжком або декількома ланцюжками амінокислот в строго певній послідовності. Ця послідовність називається первинною структурою білка, що також у значній мірі визначає всю будову білка, а отже і його біологічні властивості. Набір амінокислот також універсальний для переважної більшості живих організмів.
Експресія генів або реалізація генетичної інформації у живих клітинах (зокрема синтез білка, що кодується геном) здійснюється за допомогою двох основних матричних процесів: транскрипції (тобто синтезу мРНК на матриці ДНК) і трансляції генетичного коду в амінокислотну послідовність (синтез поліпептідного ланцюжка на матриці мРНК). Для кодування 20 амінокислот, а також стоп-сигналу, що означає кінець білкової послідовності, достатньо трьох послідовних нуклеотидів. Набір з трьох нуклеотидів називається кодоном. Прийняті скорочення, що відповідають амінокислотам і кодонам, зображені на малюнку

Генетичний код
 Комплементарні нуклеотиди
Комплементарні нуклеотиди
12. Використання ДНК в технології
З розвитком молекулярної біології було розроблено багато методів роботи з ДНК. Ці методи перш за все включають виділення ДНК, зазвичай за допомогою руйнування клітин, що містять необхідну ДНК, та спиртової преципітації ДНК з розчину. При необхідності ДНК очищують за допомогою адсорбційної хроматографії. Більші кількості ДНК можна одержати за допомогою полімеразної ланцюгової реакції (ПЛР), що вимагає лише кількох молекул ДНК, але дозволяє ампліфікувати лише відносно невеликі ділянки ДНК, зазвичай до 1500 бp, або молекулярного клонування для ділянок більшої довжини.
Отримана ДНК може бути проаналізована за допомогою рестрикційного аналізу, тобто розрізання ДНК на певних ділянках за допомогою рестриктаз, та розділення отриманих фрагментів за допомогою гелевого електрофорезу, а потім, якщо необхідно, їхньої візуалізації за допомогою саузерн-блоту. В деяких випадках можливий одночасний аналіз цілих геномів, для чого використовуються ДНК-мікрочіпи, тобто матриці, на які нанесені флюоресцентно мічені комплементарні ДНК, що дозволяє проведення порівняльної гібридизації геномів та аналіз рівня експресії бігатьох генів одночасно (хоча в останньому випадку мова йде про детекцію РНК, а не ДНК).
Ще одним з поширених методів роботи з ДНК є секвенування, тобто встановлення її нуклеотидної послідовності. Численні проекти секвенування та аналізу ДНК в останні роки 20-го століття та на початку 21-го привели до встановлення послідовностей та опису геномів багатьох організмів всіх головних таксономічних груп. Найбільшим та найвідомішим з них став проект геному людини.

(Виділення ДНК методом спиртової преципітації. ДНК виглядає як клубок білих ниток.)
13.Геннна інженерія
Сучасні біологія і біохімія інтенсивно використовують методи, засновані на рекомбінантній ДНК. Рекомбінантна ДНК — штучно створена людиною послідовність ДНК, частини якої можуть бути синтезовані хімічним шляхом, за допомогою ПЛР, або клоновані з ДНК різних організмів. Рекомбінантні ДНК можуть бути трансформовані в клітини живих організмів у складі плазмідабо вірусних векторів. Генетично модифіковані тварини і рослини зазвичай містять рекомбінантні гени, вбудовані в їхні хромосоми. Тоді як генетично модифіковані бактерії і дріжджівикористовуються для виробництва рекомбінантних білків, тварини використовуються в медичних дослідженнях, а рослини з покращеними харчовими якостями — в сільському господарстві. Судово-медична експертиза
Судмедексперти використовують знайдені на місці злочину ДНК крові, сперми, шкіри, слини або волосся для ідентифікації злочинця. Процес ідентифікації називається генетичним фінґерпринтингом або визначенням картини (профайлу) ДНК. У фінґерпринтингу порівнюється варіабельні ДНК геному, наприклад, короткі тандемні повтори й послідовності мінісателітів різних людей. Це дуже надійний метод визначення злочинців, хоча визначення може бути утруднене при забрудненні сцени злочину ДНК інших людей.
Фінґерпринтинг був розроблений в 1984 році британським генетиком Алеком Джеффрейсом і вперше використаний як доказ у суді над Коліном Пітчфорком в справі, де він був звинувачений у вбивстві й згвалтуванні.
Наразі в багатьох західних країнах, наприклад, Великобританії і США, у злочинців, звинувачених у злочинах деяких типів, забирається зразок ДНК для бази даних. Це допомогло визначити винних в раніше нерозкритих злочинах, оскільки ДНК зберігається на речових доказах. Ще цей метод використовується для визначення особи у разі масової загибелі людей та багатьох інших тестах. Зокрема, крім встановлення злочинців, метод фінґерпринтингу використовується для проведення тесту на батьківство, встановлення відповідності донорських органів, діагностики генетичних хвороб та дослідження популяцій тварин.
14.Біоінформатика
Біоінформатика включає обробку даних, що міститься в послідовності ДНК. Розвиток комп'ютерних методів зберігання і пошуку такої інформації привів до розвитку таких напрямів інформатики, що знайшли й інше застосування, як ССА, машинне навчання і організація баз даних. Алгоритми типу ССА, які шукають певну послідовність «букв» у більшій послідовності букв, були розроблені для пошуку специфічних послідовностей нуклеотидів. В інших комп'ютерних застосуваннях, наприклад, текстових редакторах найпростіші алгоритми справляються з цим завданням, але прогляд послідовності ДНК належить до складних задач, тому що вони дуже великі й складаються всього з чотирьох букв. Схожа проблема виникає при порівнянні послідовностей із різних організмів, яке використовується у вивченні філогенетичних взаємин між цими організмами й функцій білків. Дані про послідовність цілих геномів, одним з найскладнішим з яких є геном людини, важко використовувати без опису, що вказує на положення генів і регуляторних послідовностей на кожній хромосомі. Ділянки ДНК, послідовності якої містять фрагменти, асоційовані з генами, що кодують білки або РНК, можуть бути знайдені за допомогою спеціальних алгоритмів, які дозволяють передбачити наявність продуктів експресії генів до їхнього виявлення в результаті експериментів.
ДНК вперше була використана в обчислювальній техніці для розв'язку «проблеми гамільтонового шляху», окремого випадку NP-повної задачі. ДНК-комп'ютер має переваги над електронними комп'ютерами, оскільки теоретично вимагає менше енергії, займає менше місця і ефективніший завдяки можливості одночасних підрахунків. Інші задачі, наприклад, задача «абстрактних машин», задача здійсненності бульових формул і варіант задачі комівояжера були проаналізовані за допомогою ДНК-комп'ютерів. Завдяки компактності ДНК вона теоретично може знайти застосування в криптографії, де може використовуватися для конструювання одноразових шифроблокнотів.
15. Історія і антропологія
Оскільки з часом в ДНК накопичуються мутації, які потім передаються у спадок, вона містить історичну інформацію, тож генетики можуть досліджувати еволюційну історію організмів (філогенетику).
Філогенетика — метод еволюційної біології. Якщо порівнюються послідовності ДНК усередині виду, еволюційні генетики можуть довідатися історію окремих популяцій. Ця інформація може бути корисною в різних областях науки, починаючи з екологічної генетики й закінчуючи антропологією, наприклад, ДНК використовувалася для ідентифікації десяти втрачених колін ізраїльових. ДНК використовується для визначення батьківства і споріднених взаємин, наприклад, було доведено, що третій президент США Томас Джефферсон був батьком дитини рабині Салі Гемінгс. У Росії останки родини останнього царя Російської імперії Миколи II були також ідентифіковані за допомогою зразків ДНК, узятої у родичів, що живуть нині. Метод, що використовується в таких випадках, схожий на метод, який застосовують у криміналістиці.
Генетична генеалогія — набір генетичних методів, що використовуються для досліджень з генеалогії. Зазвичай генетична генеалогія використовує ДНК-тести спільно з традиційними генеалогічними методами дослідження. Успіх традиційних методів цілком залежить від збереження і існування документів (наприклад, переписних книг, літописів і т. д.). Кожна людина несе в собі свого роду «біологічний документ», який не може бути загублений, — це ДНК людини. Методи генетичної генеалогії дозволяють зокрема дістати доступ до тієї частини ДНК, яка передається незмінною від батька до сина по прямій чоловічій лінії, — Y-хромосомі.
ДНК-тест Y-хромосоми дозволяє, наприклад, двом чоловікам визначити чи розділяють вони загального предка по чоловічій лінії чи ні. ДНК-тест не просто допомога в генеалогічних дослідженнях — це сучасний інструмент, який генеалоги можуть використовувати для того, щоб встановити або спростувати споріднені зв'язки між декількома людьми.
В цьому тесті використовуються спеціальні ДНК-маркерів, нуклеотидна послідовність в яких повторюється безліч разів, — «короткі тандемні повтори». Ці повтори можна знайти і проаналізувати за допомогою таких методів як ПЛР або секвенування ДНК. Прикладом STR може бути така послідовність ДНК так:
…ЦTГT TЦTA TЦTA TЦTA TЦTA TЦTA TЦTA TЦTA TЦTA TЦTA TЦTГЦЦ…
Можна відмітити, що TЦTA повторюється 9 разів, а оскільки цей STR-маркер називається DYS391 (DNA Y-chromosome Segment № 391) робиться запис: DYS391 = 9.
У цьому маркері число повторень може бути між 7 і 14. Y-хромосома унікальна в цьому відношенні, тому що не піддається кросинговеру з кожним новим поколінням. В результаті злиття яйцеклітини і сперматозоїда, дитина отримує гени, які будуть сумішшю генів батька і матері. Але Y-хромосома передається тільки від батька, таким чином число повторів в маркерах сина буде тим же самим, що і у його батька. Діаграма нижче показує шлях Y-хромосоми, яка подорожує вниз по всіх чоловічих лініях, таким чином генетичні кузени також розділять ту ж саму Y-хромосому.
Іноді число повторів збільшується або зменшується, зазвичай в одній з ліній. Таким чином, батько може мати DYS391 = 9, а його син DYS391 = 10. Це є прикладом мутації і трапляється, коли ДНК копіюється трохи неправильно. Варто відзначити, що це природне явище. Ці мутації дуже важливі, тому що учені знають частоту їх виникнення, і таким чином можна вирахувати приблизний час, коли жив найближчий спільний предок (MRCA).
Після перевірки і об'єднання результатів декілька STR з одного геному визначається гаплотип, який може бути представлений у вигляді послідовності числа кожного маркера. Тест з 21 маркера може бути схожий на дану таблицю:
| STR Маркери Y-ДНК | ||||||||||||
| 385a | 385b | 389i | 389ii | |||||||||
| Your Haplotype |
Маркери STR записані в заголовку, а сам гаплотип в елементах таблиці. Так, наприклад, для DYS19 написано 14 повторів. Гаплотип може дати інформацію про те, звідки відбулася ваша Y-хромосома, тобто прослідкувати важ шлях предків даної людини протягом 100 тисяч років. Наприклад, Атлантичний Модальний Гаплотип (AMГ) визначений тільки шістьма маркерами, і це найзагальніший гаплотип у Західній Європі.
У базі даних YHRD кожен може порівняти його гаплотип з іншими занесеними в неї зразками. Ця база даних містить велику кількість євразійських зразків, а зараз містить ще і зразки американців і жителів східної Азії, а також ескімосів. База даних YHDR використовує до одинадцяти маркерів. Окрім цього, Y base — корисний інструмент дослідника і дозволяє додати результати своїх тестів Y-хромосоми в базу даних.
Цікавий проект-база даних гаплотипів і генеалогічних даних — Sorenson Molecular Genealogy Foundation. Після заповнення в критеріях пошуку гаплотипу, програма покаже в результатах найближчі за збігами гаплотипи з прізвищами людей і покаже генеалогічне дерево, де буде показаний передбачуваний загальний предок і всі інші зразки, з якими збіглися результати маркерів. У цій базі даних зарегістровано більше 50000 гаплотипів.
Тестування Y-хромосоми найцікавіше, якщо порівнювати результати двох і більше чоловік спільно з результатами традиційних генеалогічних пошуків. Нижче описаний гіпотетичний випадок, де три генетичні кузени з одним прізвищем пройшли тест. У якийсь момент у минулому цієї сім'ї відбулася єдина мутація в Y-хромосомі. Ця мутація залишила слід в ДНК всіх чоловіків цієї сім'ї. При порівнянні їх гаплотіпов спостерігається наступне:
| Y-ДНК STR-маркери | ||||||||||||
| 385a | 385b | 389i | 389ii | |||||||||
| Кузен 1 | ||||||||||||
| Кузен 2 | ||||||||||||
| Кузен 3 |
У цій таблиці більшість чисел збігаються, за винятком маркера поміченого сірим кольором. У учасника № 3 показана мутація в DYS392. Учасники № 1 і № 2, цифри яких повністю збігаються, дуже близькі родичі. Учасник № 3 теж є їх родичем, але дальшим.
Генетична генеалогія допомагає підтвердити результати традиційних архівних досліджень, показуючи що дві або більше людини з тим же прізвищем пов’язані спорідненістю, тобто мають загального предка. Оцінка часу життя їх гіпотетичного Спільного Предка зводиться до математики і статистики. Дослідження показують, що мутація в будь-якому маркері — окремий випадок, і відбувається приблизно кожні 500 поколінь (тобто раз в 10000 років). Якщо є точний збіг в 21 маркері, то середній час, що пройшов з тих пір, коли жив найближчий спільний предок (MRCA), тільки 8,3 поколінь. Якщо є хоч би одна єдина розбіжність (мутація), тоді час збільшується до 20,5 поколінь.
Скільки мутацій (невідповідностей) повинне бути присутнім в результатах тестів двох людей, щоб можна було виключити їх приналежність до одного клану? Велика кількість мутацій говорить про віддаленішу спорідненість або його відсутність. У випадку з 21 маркером 2 мутації між гаплотипами — це прикордонний результат, а 3 мутації зазвичай виключають взагалі достатньо близьку спорідненість між цими людьми (в межах тисячоліть).
Висновок:
ДНК служить матрицею для синтезу рибонуклеїнових кислот (РНК), визначаючи тим самим їх первинну структуру (транскрипція). Через посередництво інформаційної РНК (і-РНК) здійснюється трансляція - синтез специфічних білків, структура яких задана ДНК у вигляді певної нуклеотидної послідовності. Отже, якщо РНК переносить біологічну інформацію, «записану» в молекулах ДНК, на синтезовані молекули білків, то ДНК зберігає цю інформацію і передає її у спадок.Ця роль ДНК доводиться тим, що очищена ДНК одного штаму бактерій здатна передавати іншому штаму ознаки, характерні для штаму-донора, а також тим, що ДНК вірусу, що мешкав в прихованому стані в бактеріях одного штаму, здатна переносити ділянки ДНК цих бактерій на другий штам при зараженні його цим вірусом і відтворювати відповідні ознаки у штаму-реципієнта. Спадкові задатки (гени) матеріально втілені в певній послідовності нуклеотидів в ділянках молекули ДНК і можуть передаватися від одного індивідуума іншому разом з цими ділянками.Спадкові зміни організмів (мутації) пов'язані зі зміною, випаданням або включенням азотистих основ у полінуклеотидних ланцюжка ДНК і можуть бути викликані фізичними або хімічними впливами. З'ясування будови молекул ДНК і їх зміна - шлях до отримання спадкових змін у тварин, рослин і мікроорганізмів, а також до виправлення спадкових дефектів.
Використаналітература:
1.Альбертс Б., Брей Д., Льюис Дж. та ін Молекулярна біологія клітини в 3-х томах. - М.: Мир, 1994.
2. Докінз Р. Егоїстичний ген. - М.: Мир.
3. Історія біології з початку XX століття до наших днів. - М.: Наука, 1975.
4. Льюін Б. Гени. - М.: Мир, 1987.
5. Пташний М. Переключення генів. Регуляція генної активності і фаг лямбда. - М.:Мир, 1989. Всі форуми Книга «перемикання генів» М. Пташний
6. Уотсон Дж. Д. Подвійна спіраль: спогади про відкриття структури ДНК. - М.: Мир,1969.

