




Аллергические реакции, формирующиеся по I типу иммунного повреждения, называют атопическими (реагиновыми, анафилактическими). Их развитие характеризуется следующими особенностями:
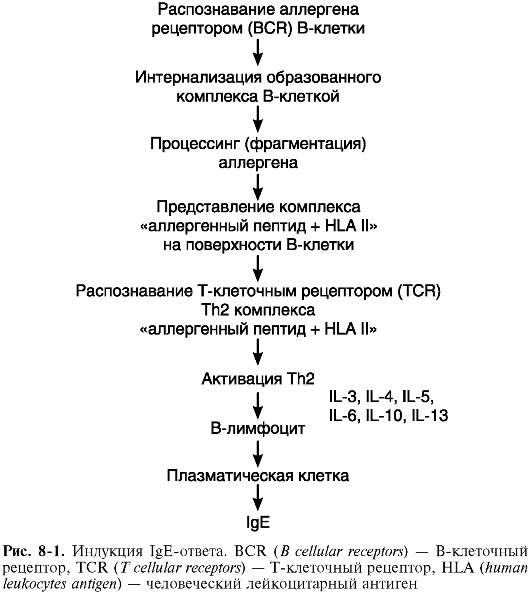
I. Стадия иммунных реакций. В норме человеческий организм толерантен к аллергенам окружающей среды. У лиц с наличием определенных молекулярных аномалий иммунной системы (в частности, при наличии очень высокой экспрессии специфических рецепторов (FceRI) на тучных клетках и базофилах крови, что определяется геном на 11-й хромосоме) при избыточном поступлении ряда веществ в организм развивается активный иммунный ответ - (сенсибилизация) к аллергенам. Этот вариант ответа является Тh2- зависимым, регулируется интерлейкином (IL) 4, IL-13, другими цитокинами и заканчивается образованием плазматических клеток, синтезирующих IgE и IgG4 (рис. 8-1). В ходе сенсибилизации происходит взаимодействие следующих клеток: дендритных (обеспечивают эндоцитоз аллергена, его процессинг и презентацию в комплексе с HLA II), Т-лимфоцитов-хелперов 2-го типа (вырабатывают цитокины соответствующего профиля, которые нужны для регуляции) и В-лимфоцитов (они сами могут выступать в качестве антигенпредставляющих, но главное - они дифференцируются в плазматические клетки-антителопродуценты и В-клетки памяти) (рис. 8-2). В-клетки памяти при повторном поступлении в организм причинно-значимого аллергена обеспечивают экспрессный (в течение 10-20 мин) ответ на него, что соответствует следующим стадиям патологического процесса. Течение сенсибилизации регулируется не только цитокинами профиля Тh2, но и соответствующими костимулирующими молекулами (CTLA-4, CD3OL, CD4OL, OX40 и др.). Второй тип рецепторов IgE - низкоспецифический FceRII (CD23) - экспрессируется на В-лимфоцитах, эозинофилах, макрофагах и тромбоцитах, обеспечивает проявления IgE-опосредованной цитотоксичности по отношению к паразитам, регуляцию синтеза IgE и ряд других функций.
 IgE значительно отличаются по своим свойствам от других антител (см. табл. 8-1). Прежде всего, они обладают цитотропностью (цитофильностью), что определяет трудность их обнаружения, так как они не участвуют в серологических реакциях. Считается, что присущее IgE свойство прикрепляться к клеткам и фиксироваться в тканях связано с приобретенными в филогенезе дополнительными 110 аминокислотами на Fс-фрагменте молекулы. Концентра-
IgE значительно отличаются по своим свойствам от других антител (см. табл. 8-1). Прежде всего, они обладают цитотропностью (цитофильностью), что определяет трудность их обнаружения, так как они не участвуют в серологических реакциях. Считается, что присущее IgE свойство прикрепляться к клеткам и фиксироваться в тканях связано с приобретенными в филогенезе дополнительными 110 аминокислотами на Fс-фрагменте молекулы. Концентра-
 Рис. 8-2. Патогенез аллергических реакций I (реагинового, анафилактического) типа
Рис. 8-2. Патогенез аллергических реакций I (реагинового, анафилактического) типа
Таблица 8-1. Биологические свойства иммуноглобулинов
 Примечание. «+» - наличие; «±» - слабая выраженность, «-» - отсутствие свойства
Примечание. «+» - наличие; «±» - слабая выраженность, «-» - отсутствие свойства
ция IgE в сыворотке крови потому и низка, что синтезируемые в региональных лимфоузлах молекулы IgE в меньшей степени попадают в кровоток, так как в основном фиксируются в окружающих тканях. Разрушение или инактивация этого участка Fс-фрагмента нагреванием (до 56 °С) приводит к потере цитотропных свойств этих антител, т.е. они термолабильны.
Фиксация антител клетками происходит при помощи рецептора, встроенного в мембрану клеток. Самой высокой способностью связывать IgE-антитела обладают рецепторы для IgE, найденные на тучных клетках и базофилах крови, поэтому эти клетки получили название клетки-мишени I порядка. На одном базофиле может фиксироваться от 3000 до 300 000 молекул IgE. Рецептор для IgE обнаружен также на макрофагах, моноцитах, эозинофилах, тромбоцитах и лимфоцитах, однако их связывающая способность ниже. Эти клетки получили название клетки-мишени II порядка (рис. 8-3).
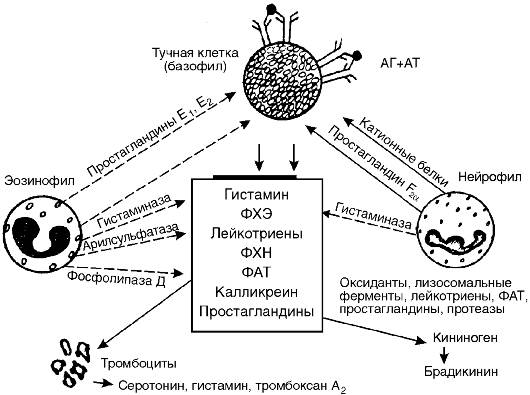 Рис. 8-3. Кооперация клеток-мишеней и взаимодействие медиаторов аллергических реакций I типа. ФХЭ - фактор хемотаксиса эозинофилов, ФХН - фактор хемотаксиса нейтрофилов, ФАТ - фактор активации тромбоцитов
Рис. 8-3. Кооперация клеток-мишеней и взаимодействие медиаторов аллергических реакций I типа. ФХЭ - фактор хемотаксиса эозинофилов, ФХН - фактор хемотаксиса нейтрофилов, ФАТ - фактор активации тромбоцитов
Связывание IgE на клетках - зависимый от времени процесс. Оптимальная сенсибилизация может наступить через 24-48 ч.
Итак, первичное попадание аллергена в организм через кооперацию дендритных клеток, Т- и В-лимфоцитов запускает сложные механизмы синтеза IgE, фиксирующихся на клетках-мишенях. Повторный контакт организма с этим аллергеном приводит к образованию комплекса АГ+АТ, связанного с поверхностью клеткимишени посредством молекул IgE. При этом условием, достаточным для активации и дегрануляции клеток-мишеней, является связывание аллергена по крайней мере с двумя соседними молекулами IgE. Начинается II стадия аллергической реакции.
II. Стадия биохимических реакций. В этой стадии основную роль играют тучные клетки и базофилы крови, т.е. клетки-мишени I порядка. Тучные клетки (тканевые базофилы) - это клетки соеди-
нительной ткани. Они обнаруживаются преимущественно в коже, дыхательных путях, по ходу кровеносных сосудов и нервных волокон. Тучные клетки имеют большие размеры (10-30 мкм) и содержат гранулы диаметром 0,2-0,5 мкм, окруженные перигранулярной мембраной. Гранулы тучных клеток и базофилов крови содержат медиаторы: гистамин, гепарин, фактор хемотаксиса эозинофилов (ФХЭ), фактор хемотаксиса нейтрофилов (ФХН) (табл. 8-2).
Таблица 8-2. Медиаторы аллергических реакций I типа

 Образование комплекса АГА+Т, фиксированного на поверхности тучной клетки (или базофила крови), приводит к стягиванию белков-рецепторов для IgE, клетка активируется и секретирует медиаторы. Максимальная активация клетки достигается связыванием нескольких сотен и даже тысяч рецепторов.
Образование комплекса АГА+Т, фиксированного на поверхности тучной клетки (или базофила крови), приводит к стягиванию белков-рецепторов для IgE, клетка активируется и секретирует медиаторы. Максимальная активация клетки достигается связыванием нескольких сотен и даже тысяч рецепторов.
В результате присоединения аллергена рецепторы приобретают энзиматическую активность и запускается каскад биохимических реакций. Активируются мембраносвязанные ферменты - фосфолипаза С и аденилатциклаза, катализирующие реакции с образованием соответственно инозитол-1,4,5-трифосфата, 1,2- диацижлицерина и цАМФ. Инозитол-1,4,5-трифосфат и цАМФ обеспечивают фосфорилирование и активацию Са2+-связывающего белка кальмодулина, мобилизующего Са2+ из эндоплазматического ретикулума клеток в цитоплазму, в присутствии которого при участии цАМФ и 1,2-диацилглицерина активируется протеинкиназа С. Протеинкиназа С осуществляет фосфорилирование и активацию ряда других внутриклеточных ферментов, в частности Са2+-зависимой фосфолипазы А2. При этом за счет Са2+- индуцированного сокращения микротрубочек гранулы «подтягиваются» к плазматической мембране, а 1,2-диацилглицерин, продукты его расщепления (моноацилглицерин, лизофосфатидиловая кислота) и активации фосфолипазы А2 (лизофосфатидилхолин) обусловливают слияние депонирующих гранул тучной клетки (или базофила крови) со стенкой мембраносвязанных канальцев и цитоплазматической мембраной, через которые медиаторы гранул (первичные) и медиаторы, образующиеся при активации клеток (вторичные; см. табл. 8-2), высвобождаются наружу. Источником
вновь образующихся медиаторов в клетках-мишенях являются продукты распада липидов: фактор активации тромбоцитов (ФАТ), простагландины, тромбоксаны и лейкотриены.
Следует отметить, что при псевдоаллергических реакциях (см. раздел 8.2) дегрануляция тучных клеток и базофилов может происходить и под влиянием неиммунологических активаторов, т.е. быть IgE-независимой.
В результате выделения из тучных клеток и базофилов факторов хемотаксиса нейтрофилов и эозинофилов последние скапливаются вокруг клеток-мишеней I порядка. Нейтрофилы и эозинофилы активируются и тоже высвобождают биологически активные вещества и ферменты. Часть из них являются также медиаторами повреждения (например, ФАТ, лейкотриены и др.), а часть (гистаминаза, арилсульфатаза, фосфолипаза D и др.) - ферментами, разрушающими определенные медиаторы повреждения. Так, арилсульфатаза эозинофилов вызывает разрушение лейкотриенов, гистаминаза - разрушение гистамина. Образующиеся простагландины группы E снижают высвобождение медиаторов из тучных клеток и базофилов.
III. Стадия клинических проявлений. В результате действия медиаторов повышается проницаемость сосудов микроциркуляторного русла, что сопровождается развитием отека и серозного воспаления. При локализации процесса на слизистых оболочках возникает гиперсекреция. В органах дыхания развивается бронхоспазм, который, наряду с отеком стенки бронхиол и гиперсекрецией мокроты, обусловливает резкое затруднение дыхания. Все эти эффекты клинически проявляются в виде приступов бронхиальной астмы, ринита, конъюнктивита, крапивницы (гиперемия и волдыри), кожного зуда, местного отека, диареи и др. В связи с тем что одним из медиаторов является ФХЭ, очень часто I тип аллергии сопровождается увеличением количества эозинофилов в крови, мокроте, серозном экссудате.
В развитии аллергических реакций I типа выделяют раннюю и позднюю стадии. Ранняя стадия появляется в течение первых 10-20 мин в виде характерных волдырей. В ней преобладает влияние первичных медиаторов, выделяемых тучными клетками и базофилами.
Поздняя стадия аллергической реакции развивается через 2-6 ч после контакта с аллергеном и в основном связана с действием вторичных медиаторов. Она характеризуется отеком, краснотой,
уплотнением кожи, которое формируется в течение 24-48 ч с последующим образованием петехий. Морфологически поздняя стадия характеризуется наличием дегранулированных тучных клеток, периваскулярной инфильтрации эозинофилами, нейтрофилами, лимфоцитами. Окончанию стадии клинических проявлений способствуют следующие обстоятельства:
а) в ходе стадии III удаляется повреждающее начало - аллерген. Активируется цитотоксическое действие макрофагов, стимулируется выделение энзимов, супероксидного радикала и других медиаторов, что очень важно для защиты против гельминтов;
б) благодаря в первую очередь ферментам эозинофилов устраняются повреждающие медиаторы аллергической реакции.

