




Высокую чувствительность к повреждающему воздействию болезнетворных факторов проявляют такие внутриклеточные структуры, как митохондрии, рибосомы и полисомы, лизосомы, элементы цитоскелета.
Митохондрии. Нарушение биоэнергетических функций митохондрий - одно из наиболее ранних проявлений повреждения клеток. Например, после прекращения кровообращения происходит нарушение окислительного фосфорилирования в митохондриях через 20-30 мин в печени и через 30-60 мин - в почках. Приблизительно в эти же сроки появляются и другие признаки повреждения клеток.
К признакам нарушений функций митохондрий относятся:
1. Снижение потребления кислорода. Уменьшение скорости потребления кислорода митохондриями, связанное с нарушением работы переносчиков электронов, наблюдается при действии
многих токсических соединений, например ионов тяжелых металлов, таких как ртуть или серебро, ряда гидрофобных соединений, различных производных углеводорода, при ПОЛ. Оно может быть также следствием набухания митохондрий и разрыва их наружной мембраны, в результате чего из митохондрий выходит цитохром С, который является одним из переносчиков электрона по дыхательной цепи.
2. Увеличение проницаемости внутренней митохондриальной мембраны. Происходит при повреждении митохондрий вследствие гипоксии или ПОЛ.
3. Снижение способности накапливать кальций. Параллельно разобщению окислительного фосфорилирования отмечается потеря способности митохондрий к накоплению ионов кальция. В присутствии избытка субстратов дыхания и при наличии кислорода и ортофосфата митохондрии печени способны накапливать в матриксе количество фосфорнокислого кальция, по массе превышающее массу митохондрий в сотни и даже в тысячу раз! Повреждение митохондрий приводит к падению разности потенциалов на митохондриальной мембране. Положительно заряженные ионы кальция, удерживаемые в матриксе электрическим полем, начинают выходить наружу из поврежденных митохондрий.
Разобщение окислительного фосфорилирования и выход кальция из митохондрий имеют самые драматические последствия для клетки (см. выше). Снижение уровня АТФ в клетке приводит к выключению ионных насосов плазмолеммы, вхождению в клетку из окружающей среды ионов натрия и кальция и выходу калия. Это обусловливает стимуляцию целого комплекса ферментных систем, активируемых ионами кальция, включая фосфолипазы, протеинкиназы, фосфатазы, протеазы, многие системы биосинтеза белков; метаболизм клетки вначале активируется, а затем дезорганизуется. Именно повреждение митохондрий является, согласно современным представлениям, тем переломным моментом, после которого изменения в клетке, вызванные повреждающим агентом, становятся необратимыми, и клетка погибает.
4. Набухание митохондрий. Весьма важным морфологическим признаком повреждения митохондрий является их набухание. Набухание митохондрий наблюдается, например, в клетках миокарда при недостаточности сердца, а также при многих инфекционных, гипоксических, токсических и других патологических процессах. Набухание митохондрий происходит при помещении клеток в ги-
потоническую среду, под влиянием ионизирующей радиации, бактериальных токсинов, при действии на клетку химических ядов и других патогенных агентов. Набухание приводит сначала к разрывам наружных мембран митохондрий, а затем - к их полному разрушению.
В опытах с изолированными митохондриями показано, что существует два типа набухания: пассивное и активное. В противоположность плазматической мембране, сравнительно хорошо проницаемой для ионов калия и хлора, внутренние мембраны митохондрий непроницаемы для ионов, за исключением Ca2+ и, возможно, ионов железа. В изотоническом растворе КС1 неповрежденные митохондрии сохраняют свой объем, несмотря на то что концентрация ионов калия и хлора внутри этих органелл существенно меньше, чем снаружи: осмотическое давление внутри создается и другими ионами, а также белками матрикса. Поэтому пассивное набухание митохондрий при отсутствии источников энергии (субстраты дыхания, кислород, АТФ) происходит только при воздействиях, которые увеличивают проницаемость мембран митохондрий одновременно для катионов и анионов (например, для К+ и С1-). К таким агентам относятся ионы тяжелых металлов (ртути, серебра, свинца). Таким же действием обладает усиление ПОЛ в мембранах митохондрий. При одновременном увеличении проницаемости для ионов калия и хлора они начинают диффундировать в митохондрии, что приводит к повышению осмотического давления, входу воды и набуханию органелл, которое называется пассивным, так как не зависит от дыхания и энергизации.
В условиях живой клетки чаще имеет место иной тип набухания - активное набухание, связанное с работой цепи переноса электронов. Повреждение митохондрий происходит под действием малых доз тяжелых металлов, активации собственной фосфолипазы в условиях гипоксии, при перекисном окислении липидов и обусловлено, прежде всего, повышением проницаемости внутренней мембраны для катионов. В присутствии источников энергии (субстраты дыхания и кислород, АТФ) на мембранах митохондрий генерируется разность потенциалов величиной около 170-180 мВ со знаком «минус» в матриксе, под действием которой ионы К+ поступают внутрь поврежденных митохондрий. Вместе с калием в матрикс поступает ортофосфат, который переносится в электронейтральной форме через внутреннюю мембрану с помощью специального белкового переносчика. Активное (т.е. связанное с
затратой энергии) накопление фосфата калия в матриксе сопровождается входом в него воды и набуханием митохондрий.
Рибосомы и полисомы. При токсических воздействиях на клетки происходит изменение конфигурации эндоплазматического ретикулума и связанных с ним рибосом. Например, при отравлении тринитротолуолом в клетках печени мембраны эндоплазматического ретикулума и расположенные на них рибосомы принимают форму различных завитков, не наблюдающихся в нормальных клетках. Синтез белков осуществляется на полисомах. Угнетение синтеза определенных белков, например синтеза гемоглобина при гипопластической анемии в клетках костного мозга, происходит на фоне уменьшения числа полисом и их распада на отдельные рибосомы.
Лизосомы. Лизосомная система, богатая ферментами, является специализированным инструментом клеток, используемым для осуществления таких важных метаболических и физиологических процессов, как катаболизм белков, глико- и липопротеидов, нуклеиновых кислот; накопление, трансформация и выведение из организма чужеродных веществ, в том числе лекарств, эндотоксинов; везикулярный транспорт и рециклизация рецепторов, ауто- и гетерофагоцитоз, апоптоз, адаптация и реконструкция клеточных структур и др. Особенно важной является роль лизосомного аппарата клетки при действии болезнетворных факторов. Лизосомы участвуют в защите клеток от бактерий, инородных тел, химических веществ, в воспалительных и иммунологических реакциях, в процессах дистрофии, некроза.
Различные повреждающие агенты, например эндотоксины бактерий брюшнотифозной группы, а также мелкие неорганические частицы (двуокись кремния, двуокись титана, алмазная пыль), попадая в лизосомы, разрушают их. При этом ферменты (гидролазы, оксидоредуктазы), заключенные в лизосомах, освобождаются в цитоплазму, активируются и вызывают повреждение субклеточных структур и макромолекул, что может привести клетку к гибели.
Однако активация лизосомальных ферментов может происходить не только под действием тех или иных специфических факторов, но и в результате ацидоза, характерного для неспецифической реакции клетки на повреждающее воздействие. Одним из процессов, вызывающих выход лизосомальных ферментов, является также активация пероксидации липидов в мембранах лизосом.
С повреждающим действием лизосомных факторов связывают развитие ряда заболеваний печени, почек, злокачественных новообразований, системной красной волчанки, ревматизма, ревматоидного артрита и др.
Увеличение численности и размеров лизосом является одной из форм структурно-функциональной адаптации клеток к повреждающему воздействию. Так, многие тяжелые металлы при поступлении в организм в течение длительного времени или в высоких концентрациях способны не только накапливаться в лизосомных везикулах различных клеток, но и индуцировать усиленное образование новых первичных лизосом и их последующее набухание. Это обеспечивает защиту клеток от повреждения путем аккумуляции и обособления токсичного металла в лизосомных везикулах, а в случае повреждения клеточных органелл - быструю ликвидацию дефектных структур с помощью лизосомного аутофагоцитоза и апоптоза.
Эффект индукции образования лизосом, морфологически выявляемый в виде увеличения их численности и размеров, продемонстрирован в гепатоцитах при циррозе печени у человека, в клетках почек и печени овец, отравленных медью, в эпителии проксимальных канальцев почек у крыс при многократном введении солей кадмия, в различных участках мозга при введении ацетата свинца крысам.
Цитоскелет. Цитоплазма клетки, помимо цитозоля и клеточных органелл, как правило, содержит еще и нитевидные белковые структуры, которые в массе формируют клеточный скелет. Цитоскелет - это система белковых нитей, заполняющих цитоплазму. Он отвечает за динамичную архитектуру клетки, ее подвижность, форму. Цитоскелет состоит из трех основных структур: микрофиламентов, микротубул и промежуточных филаментов. Каждая из них состоит из одного или двух основных белков: микрофиламенты - из актина, микротубулы - из тубулина, промежуточные филаменты - из специальных белков, различных в разных тканях (кератинов - в эпителии, десмина - в мышцах, виментина - в тканях внутренней среды: соединительная ткань, хрящ, кость и др., белков нейрофиламентов - в нейронах).
Цитоскелет - динамичное образование. Его строение может меняться за счет полимеризации и деполимеризации образующих его структур - нитей и микротубул. Благодаря такой полимеризациидеполимеризации цитоскелет непрерывно перестраивается, что
лежит в основе изменений формы, движений клеток, клеточного деления, внутриклеточного транспорта, секреции и др. К элементам цитоскелета могут присоединяться «моторные» молекулы: к актиновым микрофиламентам - молекулы миозина, к микротубулам - денеина и кинезина. При этом один конец такой молекулы прикрепляется к нити или микротубуле, а другой - к органелле или соседним элементам цитоскелета. В присутствии АТФ эти молекулярные «моторы» могут перемещать органеллы вдоль элементов цитоскелета и относительно друг друга. В нервных клетках микрофиламенты и микротубулы принимают участие в монтаже системы антероградного и ретроградного аксонального транспорта биологически активных веществ.
Как известно, белки синтезируются в перикарионе нейрона, а затем транспортируются с помощью этой системы на периферию клетки (в область синаптических контактов). Транспорт по аксону необходим и для обеспечения функций аксональной мембраны - для оборота глобулярных протеинов в мембране аксонов, выполняющих функции ионных каналов. Состояние шванновских клеток также контролируется трофическими веществами, переносимыми с аксональным током. С аксональным током перемещается весь набор ферментов, нейромедиаторов и их предшественников, необходимых для обеспечения синаптической передачи нервных импульсов.
Хорошо известно, что клетки способны переключаться с одной программы работы на другую под влиянием соответствующих сигналов: клетка может начать или прекратить деление, может дифференцироваться или включить программу самоубийства - апоптоза. В последние годы появились основания утверждать, что одним из факторов, регулирующих эти процессы, могут быть изменения натяжения цитоскелета.
При действии разнообразных альтерирующих агентов на изолированные клетки (культура ткани) выявляется отчетливое изменение формы их поверхности: появляются выпячивания цитоплазмы, называемые пузырьками (от blebs). Такое «пузырение» (или вскипание - blebbing) клеточной мембраны - один из ранних и надежных признаков разрушения сети цитоскелета. «Вскипание» мембраны инициируют вещества, нарушающие гомеостаз внутриклеточного кальция.
Механизм феномена объясняют следующим образом. Кальций вовлечен в процесс поддержания структуры цитоскелета как не-
посредственно, так и через ряд Са2+-связьгаающих белков и Са2+- зависимых ферментов. Особенно значим уровень Са2+ в цитоплазме для образования ассоциации белков цитоскелета с белками плазматической мембраны и взаимодействия различных элементов цитоскелета. Стойкое увеличение концентрации кальция в цитозоле, по-видимому, вызывает отщепление актина микрофиламентов от актинина - белка, служащего промежуточным звеном, который связывает микрофиламенты цитоскелета с белками плазматической мембраны.
Кроме того, Са2+ активирует протеазы, которые могут расщеплять актинсвязывающие белки, разрушая тем самым места прикрепления филаментов цитоскелета к клеточной мембране. Отщепление цитоскелета от мембраны приводит к ослаблению фиксации последней и ее «вскипанию», что и обнаруживается при действии на клетки различных токсинов (рис. 3-9). Вещества, связывающие внутриклеточныгй кальций и ингибиторы Са2+-зависимых протеаз, препятствуют «вскипанию» плазматической мембраны клеток, обработанных токсическими агентами.
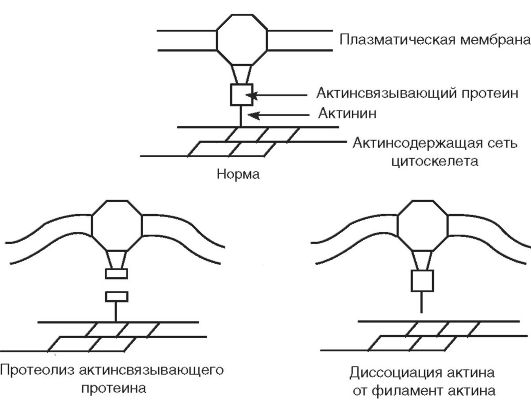 Рис. 3-9. Са2+-зависимые механизмы повреждения цитоскелета клеток, обработанных хинонами
Рис. 3-9. Са2+-зависимые механизмы повреждения цитоскелета клеток, обработанных хинонами
Другой механизм изменения конфигурации плазматической мембраны под влиянием избыточной концентрации ионов кальция может состоять в истощении внутриклеточных запасов макроэргов.
Белки цитоскелета повреждаются не только в результате повышения содержания Са2+ в цитоплазме, но и под влиянием факторов с иными механизмами действия. Так, непосредственно взаимодействуют с белками цитохалазины, фаллоидин (один из токсинов бледной поганки), а также колхицин, алкалоиды барвинка (цитостатики винбластин, винкристин).
Действие повреждающих веществ на тубулин митотического веретена делящихся клеток приводит к нарушению пролиферации последних, особенно клеток системы крови. Колхицин и алкалоиды барвинка в эксперименте, разрушая цитоскелет аксонов нервных клеток, существенно нарушают аксональный ток. Вероятно, аналогичные эффекты могут развиваться и при повреждении цитоскелета веществами, нарушающими внутриклеточный гомеостаз кальция.

