




¬ тех случа€х, когда ацетат служит не только источником энергии, но и источником различных промежуточных продуктов, необходимых дл€ син≠теза углеродных скелетов всех важных клеточных компонентов (именно так может обсто€ть дело у микроорганизмов, например у ≈. coli и Pseudomonas, у водорослей и у высших растений), цикл трикарбоновых кислот оказы≠ваетс€ заменен глиоксилатным циклом. ќбща€ схема глиоксилатного цикла представлена на рисунке 6. Ђ“опливомї дл€ этого цикла служит также ацетил- ој. ќн взаимодействует с оксалоацетатом, в результате чего, в конеч≠ном счете, образуетс€ изоцитрат. ќднако дальнейшее превращение осуще≠ствл€етс€ не обычным способом, т. е. под действием Ќјƒ-зависимой изоцитратдегидрогеназы, а путем альдольного расщеплени€, катализируемого ферментом изоцитратазои, продуктами которого €вл€ютс€ сукцинат и глиоксилат.
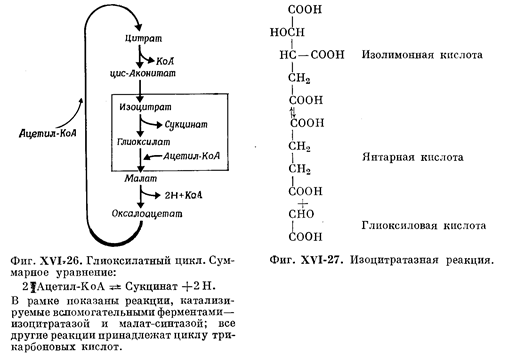
ќбразовавшийс€ глиоксилат соедин€етс€ с другой молекулой ацетил- ој. Ёта реакци€, катализируема€ ферментом малат-синтазой, приводит к образованию малата. «атем малат окисл€етс€ до оксалоацетата, который вновь мо≠жет взаимодействовать с молекулой ацетил- ој. ѕри каждом обороте глиоксилатного цикла две молекулы ацетил- ој включаютс€ в цикл, образуетс€ одна молекула сукцината Ч этот сук≠цинат используетс€ затем в процессах биосинтеза Ч и одна пара атомов водо≠рода переноситс€ от малата к кисло≠роду через дыхательную цепь, резуль≠татом чего €вл€етс€ окислительное фосфорилирование јƒ‘.
“аким образом, глиоксилатный цикл поставл€ет дл€ различных процессов биосинтеза как энергию, так и четырех-углеродные промежуточные продукты. ” высших животных этот метаболи≠ческий путь отсутствует; они в нем не нуждаютс€ просто потому, что двууглеродные соединени€ практически никогда не служат у них единствен≠ным клеточным топливом. ¬ семенах высших растений этот путь, напро≠тив, играет весьма важную роль, так как из ацетильных групп (источником которых служат жирные кислоты, вход€щие в состав запасных триглицеридов) таким путем, через сукцинат, образуютс€ углеводы. ’от€ реакции цикла трикарбоновых кислот протекают у высших расте≠ний в митохондри€х, ферменты глиоксилатного цикла Ч в частности, изо-цитратаза и малат-синтаза Ч локализуютс€ у них в других цитоплазматических органеллах, так называемых глиоксисомах. ¬ этих органеллах, снаб≠женных мембраной, отсутствует больша€ часть ферментов цикла трикарбо≠новых кислот и цитохромна€ система. √лиоксисомы найдены только в расти≠тельных клетках, способных превращать жирные кислоты в углеводы. ѕоскольку глиоксисомы богаты каталазой, можно думать, что они имеют общее происхождение с микротелъцами (которые называютс€ также пер-оксисомалш) или происход€т непосредственно от этих последних.
–еакции пути начинаютс€ с фосфорелировани€ глюкозы с образованием глюкозо-6-фосфата, затем следует окисление его до глюконо-1,5-лактон-6-фосфата и гидролиз последнего с образованием 6-фосфоглюконата. ƒегидратаци€ 6-фосфоглюконата по —2- и —3- атомам приводит к образованию енола, который спонтанно и практически необратимо таутомеризуетс€ в 2-дегидро-3-дезокси-6-фосфоглюконат, называемый также 2-кето-3-дезокси-6-фосфоглюконат ( ƒ‘√). ¬ последующей альдолазной реакции формируетс€ шиффово основание между определенным остатком лизина в составе фермента и карбонильной группы субстрата, что облегчает расщепление св€зи между —3- и —4-атомами, в результате которого образуетс€ пируват и глицеральдегид-3-фосфат.
|
|
|
ќбща€ стехиометри€ этой последовательности реакций может быть выражена следующим уравнением:
 √люкоза + ј“‘ + NADP+ глицеральдегид-3-фосфат +
√люкоза + ј“‘ + NADP+ глицеральдегид-3-фосфат +
пировиноградна€ кислота + јƒ‘ + NADPH + H+
≈сли учесть, что последующее превращение глицеральдегид-3-фосфата идет по той же схеме, что и в Ёћѕ-пути и сопровождаетс€ фосфорелированием двух молекул јƒ‘, то суммарный энергетический выход пути Ёнтнера-ƒудорова составл€ет 1 молекула ј“‘ на каждый моль утилизированной глюкозы.
3. лючевые точки пересечени€ и разветвлени€ метаболических путей в клетках бактерий.
јнализ путей метаболизма как сети направленных реакций позвол€ет вы€вить ключевые элементы их организации. »сходные и конечные вещества нераз≠ветвленного пути представл€ют собой или исходные питатель≠ные вещества, или конечные продукты метаболизма (которые в свою очередь могут задерживатьс€ в клетке или выдел€тьс€ в среду), или, наконец, исходные вещества дл€ других путей ме≠таболизма, т. е. точки разветвлени€. ќбычно перва€ после точ≠ки разветвлени€ метаболического пути необратима€ (т. е. име≠юща€ существенное по абсолютной величине отрицательное Δ G' при внутриклеточных услови€х) реакци€ €вл€етс€ аллостерически регулируемой стадией. ¬ этом случае посредством ре≠гул€ции поступлени€ исходных веществ клетка распредел€ет метаболиты, используемые одновременно дл€ нескольких це≠лей. —о структурной точки зрени€ эти метаболиты представл€≠ют собой исходные точки или узловые пункты метаболической ветви. ќтсюда следует, что точки разветвлени€ и модулируе≠мые необратимые реакции €вл€ютс€ теми критическими эле≠ментами, которые определ€ют все стороны химической актив≠ности клетки.
¬се типы метаболической активности клетки завис€т от рас≠пределени€ потоков трех основных реагентов: глюкозо-6-фосфата, пирувата и ацетил-—ој. Ќа рисунке схематично изображе≠ны пересечени€ различных путей метаболизма, в которых эти реагенты €вл€ютс€ исходными или конечными веществами. ќбъем потоков по каждому из путей регулируетс€ клеткой в за≠висимости от метаболических потребностей и ее состава в дан≠ный момент.
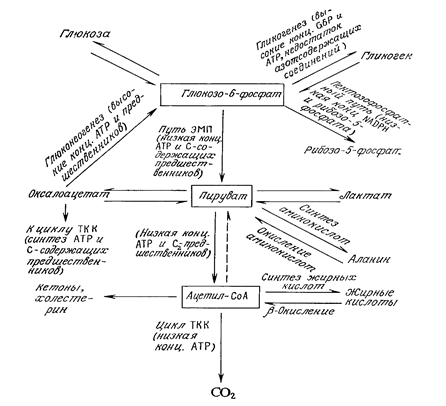
–исунок - —хема пересечени€ основных путей метаболизма углерода. ¬ клет≠ках млекопитающих превращение ацетил-—ој в пируват не происходит.
4. —ветова€ фаза фотосинтеза (аккумулирование световой энергии, транспорт электронов, фотофосфорелирование).
¬ большинстве типов дыхани€ атомы водорода посто€нно перенос€тс€ от топлива к кислороду; этот процесс сопровожда≠етс€ выделением энергии. ‘отосинтез представл€ет собой, по сути дела, обратный процесс, в котором осуществл€етс€ улав≠ливание световой энергии, используемой дл€ превращени€ ди≠оксида углерода в глюкозу и ее полимеры. ‘отосинтез €вл€ет≠с€ первичным поставщиком энергии дл€ всей биосферы. ¬ фо≠тосинтезе усваиваетс€ энерги€ единственного значимого источника энергии в —олнечной системе Ч самого солнца. ƒру≠га€ жизненно важна€ функци€ фотосинтеза заключаетс€ в его участии в круговороте углерода и кислорода; именно благодар€ фотосинтезу осуществл€етс€ восстановление углерода, окислен≠ного в ходе дыхани€:
‘отосинтез: 6—ќ2 + 6Ќ2ќ + свет Ч > C6H12O6 + 6O2

