




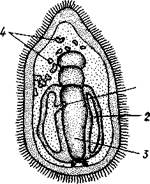
же представлена околоротовым кольцом и 5 парными нервными тяжами в руках (рис. 543). Аборальная система развита очень сильно и состоит из аборальной нервной массы, залегающей внутри особого пятикамерно-го синуса, расположенного в верхней пластинке стебелька. От нее идут 5 радиальных нервов, пронизывающих брахиальные пластинки рук и дающих ответвления в пиннулы. Специализированных органов чувств у морских лилий нет.
Особые дыхательная и выделительная системы отсутствуют.
Кровеносная система морских лилий состоит из околоротового кольца, представляющего сплетение кровеносных лакун. Оно называется губчатым органом и, возможно, выполняет роль лимфатической железы. От кольца отходят радиальные лакуны полового тяжа рук и пиннул (см. ниже). В стенках кишечника и чашечки также развиты кровеносные лакуны.
|
Перигемальная система развита очень слабо и представлена кольцом (иногда может отсутствовать) и 5 радиальными каналами в руках. Целом сильно редуцирован вследствие того, что он частично зарастает рыхлой тканью (целенхимой). Совершенно обособленный отдел целома представляет собой пятикамерный синус. Это выстланная перитонеальным эпителием полость, залегающая внутри центральной пластинки, лежащей на аборальном полюсе чашечки, и поделенная радиальными перегородками на 5 участков (рис. 543).
Половая система лилий своеобразно устроена. Crinoidea раздельнополы. От 5-камерного синуса кверху через целом чашечки поднимается губчатый лимфоид-
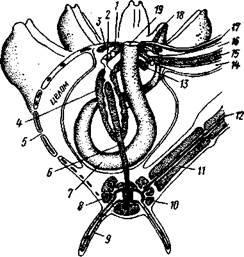
| Рис. 543. Общая анатомия морской лилии (из Никольса): U— околоротовое кольцо амбулакральной системы, 2 — околоротовое кольцо кровеносной системы, 3 — гипоневральное кольцо, 4 — осевой синус, 5 — водяная пора, 6 — осевой орган, 7 — кишечник, 8 — скелет чашечки, 9 — усик, 10 — абораль-ная нервная система в пятикамерном синусе, // — скелет руки, 12 — аборальный нервный тяж, 13 — перигемальное кольцо, 14 — кровеносный сосуд в руке, 15 — перигемальный сосуд в руке, 1€ — амбулакральный сосуд в руке, 17 — гипонев-ральный тяж в руке, 18 — анальный конус, 19 — рот |
ныи осевой орган, оральный конец которого образует половой столон. От столона в руки направляются 5 плотных клеточных половых тяжей, ветвящихся соответственно ветвлению рук. Концевые веточки тяжей заходят в пиннулы, становятся здесь полыми и превращаются в слепо замкнутые половые мешки. Клетки полового столона и его тяжей бесплодны; напротив, клетки стенок половых мешков внутри пин-нул развиваются в яйца или живчики, так что каждая пиннула содержит как бы особую половую железу. Выход половых продуктов из пиннул наружу с&вершает-ся через прорыв стенок пиннул на особо предназначенных для этого местах.
Палеонтология и систематика. Остатки представителей класса Crinoidea начинают встречаться с кембрия, причем некоторые отряды их из многих ископаемых ограничены отложениями палеозойской эры. Отложения мезозойской эры тоже содержат богатую фауну морских лилий. Ископаемые формы встречались, по-видимому, на различных глубинах и нередко достигали крупных размеров — со стебельком до 2 м длины. Известно более 300 ископаемых родов. Начиная с юрского периода морские лилии обнаруживают тенденцию отрываться от стебелька, и в срвременной фауне, относящейся к отряду Arliculata (членистые лилии), имеется всего 70 стебельчатых и около 600 бесстебельчатых лилий. Общее число видов класса Crinoidea (вместе с ископаемыми) — несколько тысяч. Из стебельчатых морских лилий в настоящее время сохранилось всего около десятка родов, причем большинство встречается лишь на больших глубинах. Одна небольшая форма, Rhizocrinus lafotensis, попадается в глубоких фиордах Норвегии.
Кроме того, насчитывается свыше десятка богатых видами родов бесстебельчатых лилий, среди которых наиболее распространена Heliometra (Средиземное море, Атлантический и Тихий океаны). У таких форм
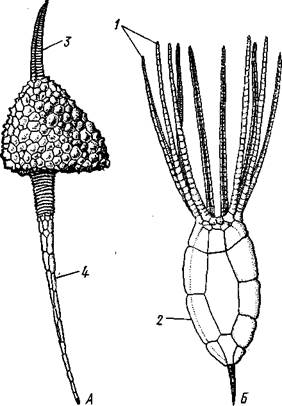
Рис. 544. Carpoidea. A—Heckericystis (ордовик); Б — Rhipidocystis (ордовик):
/ — руки, 2 — чашечка, 3 — анальный конус, 4 — стебелек
(см. рис. 541) в молодости имеется стебелек, но затем он обламывается у вершины, так что последний членик стебелька со своим венчиком усиков остается при чашечке и образует ее замыкающую центральную пластинку аборального полюса. Не-liometra держится на грунте усиками центральной пластинки, т. е. сохраняет положение своих стебельчатых предков.
КЛАСС II. КАРПОИДЕИ (CARPOIDEA)
Самая древняя вымершая группа прикрепленных или просто лежащих на субстрате иглокожих с еще не выработавшейся пятилуче-вой симметрией: тело более или менее билатерально, покрыто пластинками. Имелся небольшой полый стебелек. Известны лишь из самых древних отложений: кембрия, силура и нижнего девона.
Представители: Dendro-cyslides, Rhipidocystis (рис. 544) и др.
КЛАСС III. ЭДРИОАСТЕРОИДЕИ (EDRIOASTEROIDEA)
Вымершие, лишенные стебелька, прикрепляющиеся к субстрату или свободноподвижные пельматозои. Форма тела дисковидная или более или менее шарообразная. Многочисленные пластинки покрывали все тело. На оральной стороне располагались: в центре рот, эксцентрично анус, а также мадрепоровое отверстие. От рта по телу животного проходили 5 прямых или изогнутых радиальных амбулакральных бороздок, расположенных между двумя рядами пластинок. Между пластинками каждого ряда имелись отверстия для амбулакральных ножек. По-видимому, амбулакральная система представителей кл. Edrioasteroidea состояла из пяти радиальных каналов и соединяющего их околоротового кольца. Амбулакральные бороздки были покрыты парными кроющими пластинками (рис. 545). Возможно, что некоторые формы могли передвигаться при помощи амбулакральных ножек. Класс Edrioasteroidea очень интересен в филогенетическом отношении, так как это единственная группа среди подтипа Pelmatozoa, у которого амбулакральная система близка к таковой подтипа Eleutherozoa. С другой стороны, низшие представители класса Edrioasteroidea еще очень близки к классу Cystoidea.
Эдриоастероидеи известны с кембрия до нижнего карбона.
|
Представители: Cytaster, Ed-rioaster и др.
КЛАСС IV. ШАРОВИКИ (CYSTOIDEA)
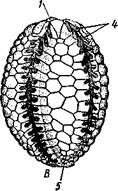
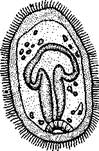
Шаровики жили исключительно в палеозое, преимущественно в силуре, и относятся к древнейшим иглокожим. Мешковидное или шаровидное тело их прикреплялось к субстрату при помощи зачаточного стебелька (рис.546). На противоположном (оральном) полюсе различимы 4 отверстия: ротовое, амбулакральное, половое и анальное (рис. 546). Тело покрыто многочисленными неправильными пластинками. Шаровики — иглокожие с асимметричным строением или неустановившейся 5-лучевой симметрией. Обычно были развиты амбулакральные желобки, расходящиеся от рта по поверхности тела радиально и продолжающиеся на небольшие возвышающиеся над телом кожные выросты — брахиоли. Желобки, вероятно, служили для собирания пищевых частиц из толщи воды. Размеры шаровиков варьировали от величины лесного ореха до величины яблока.
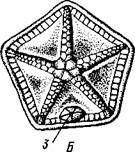
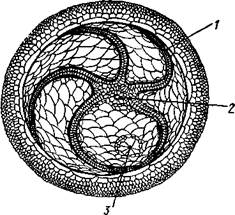
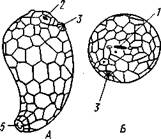
Рис. 545. Edrioasteroidea. A — Lepidodis-cus (девон); Б — Cyathocystis (ордовик):
1 — амбулакральные пластинки, 2 —мадрепо-ровая пластинка, 3 — анальная пирамида
|
|
Рис. 546. Cystoidea. A—Aristocystis (силур), ви'д сбоку; Б — то же, с орального полюса; В — Proteroblastus (силур) (по Иекелю):
/ — рот, 2 — половая пора, 3 — анальный конус, 4 — амбулакральная борозда с брахиолями, 5 — место прикрепления к субстрату
Представители: Aristocystis bohemicus (рис. 546) из нижнего силура Чехии, Sphaeronis globulus (нижний силур), Proteroblastus (рис. 546) (нижний силур), Echinosphaerites aurantium, часто попадающийся в силурийских отложениях, и др.
КЛАСС V. МОРСКИЕ БУТОНЫ (BLASTOIDEA)
Палеозойские, стебельчатые или, значительно реже, лишенные стебелька Pelmatozoa с чашевидным телом, покрытым немногочисленными правильными пластинками. В центре оральной стороны лежал рот, от которого по телу шли 5 амбулакральных бороздок, прикрытых мелкими подвижными пластинками. По бокам бороздок располагалось большое количество членистых рук (рис. 547). Морские бутоны появились в нижнем силуре и вымерли в Перми. Представители: Blaslocrinus, Pentremi-tes, Cadaster, Orophocrinus и др.
Эмбриональное развитие jEchinodermata
Развитие иглокожих на начальных стадиях идет более или менее
|
|
Рис. 547. Blastoidea. Orophocrinus (карбон) (из Безера): / — руки, 2 —чашечка с пластинками, 3 — стебель
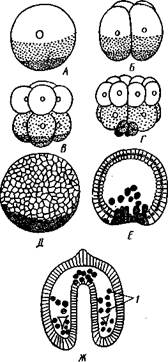
Рис. 548. Дробление яйца морского ежа Strongylocentrotus (по Давыдову). А — яйцо до начала дробления; Б — стадия четырех бластомеров; В — стадия восьми бластомеров; Г — стадия шестнадцати бластомеров; Д — бластула; Е — полярная иммиграция мезенхимы; Ж — гаструла: / — мезенхима
сходно у различных классов и всегда характеризуется сложным метаморфозом с билатеральной личиночной стадией.
|
|
| Рис. 549. Две стадии развития личинки морской звезды; энтероцельное образование целома (по Мечникову): / — зачаток каменистого канала, 2 — правый целомиче-ский мешок, 3 — кишечник личинки, 4 — клетки мезенхимы |
Дробление яйца (рис. 548) полное, радиального типа, чаще равномерное, но у морских ежей неравномерное, что связано с рано проявляющейся детерминацией. В данном случае уже на стадии 16 клеток можно различить 8 бластомеров средней величины, дающих впоследствии эктодерму животного, 4 крупных энтодермальных бла-стомера и 4 маленькие клетки, дающие мезенхиму, т. е. соединительную ткань развивающейся личинки. Дробление у всех иглокожих приводит к образованию покрытой жгутиками типичной бластулы. На нижнем полюсе последней образуется глубокое впячивание — зачаток энтодермальной средней кишки, и бластула превращается в гаструлу. Бластопор лежит на заднем конце личинки. Третий пласт — мезодерма — формируется из двух зачатков. Во-первых, производные мелких клеток уже
в самом начале образования гастрального впячивания отделяются от прочих клеток, уходят в бластоцель и дают там рыхлое скопление клеток личиночной мезенхимы. Основная же часть мезодермы взрослого животного берет начало от энтодермальной средней кишки гаструлы (рис. 549). От слепого верхнего конца кишечника отшнуровывается часть в виде самостоятельного замкнутого пузырька — зачатка целома.
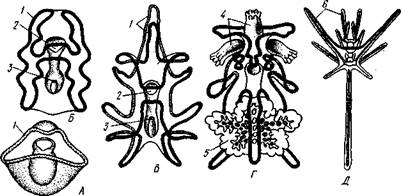
Рис. 550. Личинки иглокожих. А — диплеврула; Б — аурикулярия голотурий; В — бипиннария морской звезды; Г — брахиолярия морской звезды с формирующейся маленькой звездочкой; Д — эхиноплутеус
морского ежа:
1 — ресничный шнур, 2 — рот, 3 — анус, 4 — брахиоли, 5 — маленькая звездочка, 6 — руки
Он делится затем на два целомических мешочка, которые ложатся по сторонам кишечника. Такой способ образования целома — отделение его от кишечника — называется энтероцельным. Между тем зачаток кишечника нагибается слепым передним концом в одну сторону, которая и определяется как брюшная; навстречу ему на поверхности тела гастру-лы образуется впячивание эктодермы — зачаток передней кишки. Это впячивание сливается с загибом средней кишки. Так образуется полный кишечник. Бластопор превращается у иглокожих в порошицу (или последняя возникает на его месте), а рот образуется, как вы видели, из отдельного вторичного впячивания — признак принадлежности к группе Deuterostomia.
Далее происходит смещение порошицы на брюшную сторону, и личинка становится вполне билатерально симметричной, с выпуклой спинной и седловидно вдавленной брюшной стороной. Реснички сохраняются только на валике, окружающем рот, образуя околоротовой венчик (мерцательный шнур). Такая общая для большинства Echinodermata стадия называется диплеврулой (рис. 550, А). У разных классов диплеврула испытывает неодинаковые изменения, особенно резко проявляющиеся в видоизменении внешней формы тела и расположении околоротового мерцательного шнура.
Меньше всего изменяется форма диплеврулы у голотурий, где последняя превращается в так называемую аурикулярию. Тело аурикуля-
|
рии овальное, рот лежит в глубине брюшного вдавления, сплошной, цельный мерцательный шнур огибает вдавление, образуя выросты вперед и назад и многочисленные изгибы по направлению к спинной стороне.
Точно так же один ресничный шнур у личинок морских ежей и офиур, 'которые столь сходны, что обозначаются общим термином плутеус (рис. 550). Мерцательный шнур у плутеуса окаймляет края 4 пар длинных рук, направленных вперед. Каждая рука имеет скелет в виде известковой иголочки, залегающей по ее оси. Руки увеличивают размер, а отсюда и двигательную силу одевающего их ресничного шнура, помогая личинке держаться в планктоне и не тонуть.
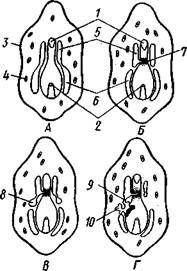
| Рис. 551. Схема развития целома у личинки иглокожих: А — Г — пос- ледовательные стадии (из Доге- ля): / — рот, 2 — анус, 3 — стенка тела, 4 — клетки мезенхимы в первичной полости тела, 5, 6 — передний и задний отделы зачатков целома, 7 — поровый канал, 8 — обособление средних отделов целома путем отшнуровывания от передних, 9 — зачаток каменистого канала, 10 — превращение левого среднего целома, гидроцеля, в кольце амбулакральной системы, правый передний и средний зачатки целома отстают в развитии и редуцируются |
У личинки морских звезд, бипиннарии (рис. 550), околоротовой венчик перешнуровывается на два замыкающихся в кольца участка — преоральный и более развитой посторальный. На более поздних стадиях ресничный шнур бипиннарии образует сложные боковые загибы, на переднем конце тела вырастают три особых отростка — брахиоли, иногда не совсем точно называемые «руками», и би-пиннария становится брахиолярией.
Как видно из описаний, личинки всех Echinodermata сначала вполне билатеральны. Закладка дефинитивного 5-луче-вого строения начинается с образо-
| 01 СЛ |
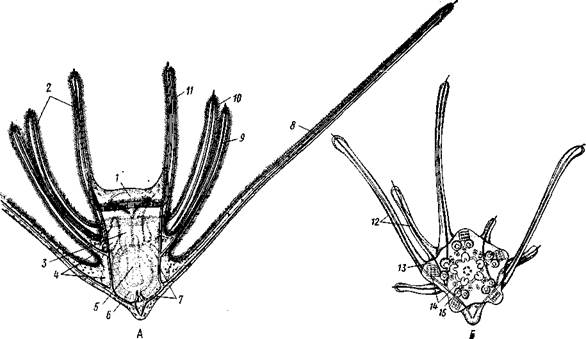
Рис. 552. Личинка и метаморфоз офиур (по Иванову). Л—личинка офиоплутеус со спинной стороны: Д —
развитие молодой офнуры в плутеусе:
/ — просвечивающий рот, 2 — ресничный шнур, 3 — передняя кишка, 4 — целомы, 5 — задняя кишка, 6 — средняя кишка, 7 — скелет, 8—11 — руки, 12 — руки личинки, 13 — тело молодой офиуры, 14 — закладывающиеся скелетные пластинки,
/5 — амбулакоальные щупальца
вания амбулакральной системы (рис. 551), чему предшествуют дальнейшие изменения целомических мешков. Именно каждый из них перешнуровывается в двух местах, давая справа и слева от кишечника уже три пары целомических мешков, свидетельствующих о трехсег-ментном (олигомерном) плане строения личинки. Оба задних (третьих) мешка преобразуются в целом взрослого животного. Первый и второй правые мешки чаще всего бывают недоразвитыми, и основные изменения, следовательно, претерпевают 2 левых передних целомических мешка, между которыми остается связь в виде канальца, превращающегося в дальнейшем в каменистый канал амбулакральной системы. При этом первый целомический мешок получает сообщение с наружной средой особым канальцем с отверстием — будущим мадрепоритом; сам же первый целомический мешок входит позже в состав осевого органа взрослого иглокожего. По мере метаморфоза средний левый целомический мешок, называемый теперь гидроцелем, т. е. зачаток всей амбулакральной системы, вытягивается и охватывает переднюю кишку личинки сначала в виде подковы с 5 слепыми выростами, а затем подкова смыкается в околоротовое кольцо; слепые выросты вытягиваются и дают начало радиальным амбулакральным каналам. Следовательно, всю амбулакральную систему можно толковать как видоизмененный целомический мешок, а каменистый канал — как целомодукт этого мешка, приобретший новую функцию.
За счет целомических мешков личинки образуется также псевдоге-мальная система взрослых иглокожих.
|
В дальнейшем метаморфоз личинки у различных классов варьирует. У голотурий извилистые выступы тела сглаживаются, ресничное кольцо распадется на 5 колец. Получающаяся бочонковидная личинка, опоясанная кольцами, словно обручами, называется куколкой. Амбулакральное кольцо ее дает 5 выпячиваний вперед, образующих первые околоротовые щупальца, и 5 выростов назад — зачатки радиальных каналов. Куколка падает на дно, теряет реснички, ложится на одну сторону и переходит к ползающему образу жизни, развивая амбулакральные ножки, и личинка целиком превращается в молодую голотурию.
У морских ежей и офиур (рис. 552) взрослое животное образуется за счет центральной области тела личинки, остальная часть которого, подобно пилидию немертин (с. 206), не принимает участия в развитии и погибает. На левой стороне плутеуса образуется глубокое впячивание эктодермы. На самом его дне эктодерма утолщается, превращаясь в зародышевый диск будущего животного.
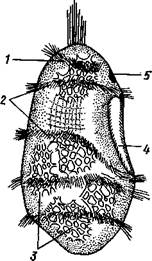
| Рис. 533. Личинка морской лилии Antedon rosacea (из Коршельта и Гейдера): I — передний ресничный поясок, прерванный на брюшной стороне, 2 — сплошные ресничные пояски, 3 — скелетные пластинки. 4 — преддверие, 5 — прикрепительная ямка |
Гидроцель располагается под диском и дает в него 5 выпячиваний, из которых образуются первые ножки ежа. Заложившееся молодое животное выпячивается наружу, руки и скелет личинки отваливаются, и молодой еж (офиура) опускается на дно. По такому же образцу, но с некоторыми изменениями, идет развитие и морских звезд.
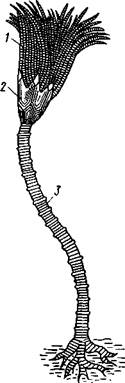
Развитие морских лилий (рис. 553; 554) хорошо известно лишь для бесстебельчатых
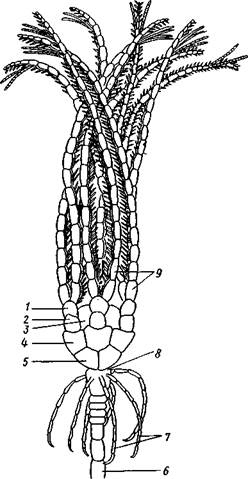
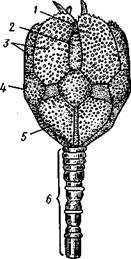
форм. Из яйца выходит личинка, похожая на бочонковидную стадию голотурий (куколку), но с теменным султаном ресниц и без рта и порошицы. После периода свободноплавающей жизни она прикрепляется ко дну передним концом. Последний утончается и превращается в стебелек, тогда как свободный (бывший задний) конец личинки булавовидно надувается и принимает форму чашечки. В мезодерме стебелька и чашечки закладываются скелетные пластинки. Сидячая стадия личинки называется пентакринусовой вследствие сходства со стебельчатыми лилиями рода Pentacrinus.
По краям чашечки вырастают выпячивания, преобразующиеся в руки. К этому времени личинка отламывается от стебелька так, что верхний членик стебелька становится центральной пластинкой (от нее вырастают усики) на аборальной стороне чашечки. Этим метаморфоз заканчивается.
|
|
Рис. 554. Пентакринусовая личинка бесстебельчатой морской лилии Heliomet-ra glacialis. А — молодая стадия; Б — поздняя стадия (по Иванову):

