




Согласно наиболее распространенному взгляду, кольчатые черви произошли от низших несегментированных червей. Признаки становления метамерии в виде правильного повторения вдоль оси тела некоторых органов, наблюдающиеся у ряда турбеллярий и немертин, а также сходство трохофоры с мюллеровской личинкой многоветвистых турбеллярий (с. 157) и пилидием немертин (с. 206) подтверждают, что тип кольчатых червей сформировался именно таким образом. Характерными признаками высокой организации кольчатых червей являются наличие у них целома и кровеносной системы. Относительно происхождения целома были высказаны различные теории. Главные из них следующие:
1. Схизоцельная ' теория утверждает, что целом кольчатых червей гомологичен первичной полости тела представителей типа Ncmathcl-minthes и представляет собой простое ее усовершенствование. Эта теория не может объяснить, почему при эмбриональном развитии у многих животных целом формируется за счет стенок кишечника.
1 Схизоцслем иногда называют первичную полость тела, возникающую в результате расхождения или распада клеток внутренних тканей. Первичная полость тела нематод и других представителей типа Nemathelminthes — типичный схизоцель.
2. Миоцельная теория — разновидность предыдущей теории. По этой теории целом образовался как полость в мускульном зачатке животного, которая заполнилась жидкостью. Целом имел сначала чисто опорное значение для окружающей его мускулатуры. Положительная сторона этой теории заключается в признании важности опорной функции цело-ма. Миоцельная теория, однако, не объясняет других функций целома.
3. Согласно гоноцельной теории целом возник из половых желез низших червей. Полость каждого целомического мешка соответствует полости гонады турбеллярии или немертины, перитонеальный эпителий происходит из стенки половой железы, а целомодукты — из половых протоков. Действительно, у всех целомических животных целом всегда несет половую функцию. Однако выведение стенки целома из стенки гонады является явной натяжкой, ибо тогда следует допустить, что часть половых клеток стала простыми соматическими клетками.
4. По энтероцельной теории целом берет начало от гастроваскулярной системы кишечнополостных и гребневиков, которые рассматриваются как отдаленные предки целомических животных. Так как у высших кишечнополостных под энтодермой гастроваскулярных каналов развиваются гонады, то энтероцельная теория хорошо объясняет половую функцию целома. Она, по существу, включает в себя и гоноцельную теорию. Кишечные поры гребневиков (с. 144) и целомодукты вторичнополостных животных, с точки зрения энтероцельной теории, могут считаться очень близкими образованиями.
Энтероцельная теория подтверждается способом формирования целома во время развития вторичноротых — Deuterostomia, у которых це-ломические мешки возникают как боковые выпячивания энтодермаль-ного кишечника, т. е. энтероцельно (с. 555). Таким образом, во всяком случае в отношении вторичноротых животных, энтероцельная теория не вызывает сомнений.
У Protostomia, однако, как мы видели, имеет место иной способ формирования мезодермы и целома. Последний во время эмбрионального развития образуется путем расхождения клеток мезодермальных полосок, а сами полоски формируются из двух первичных мезодермальных клеток (потомков бластомера 4d).
На первый взгляд, различия между телобластическим и энтероцель-ным способами закладки и формирования целома столь велики, что можно допустить независимое происхождение последнего у Protostomia и Deuterostomia. Однако необходимо помнить, что бластомер 4of принадлежит к четвертому квартету, все остальные клетки которого идут на построение энтодермальной средней кишки. Можно поэтому допустить, что бластомер 4d первоначально также принадлежал энтодерме и что мезодермальные полоски сначала были разрастаниями энтодермы. Затем уже вторично, как упрощение самого способа эмбрионального развития, они стали развиваться только из двух клеток, рано отделяющихся от энтодермы. С этой точки зрения между телобластическим способом развития целома (у первичноротых) и энтероцельным (у вторичноротых, а также плеченогих и щетинкочелюстных — с. 516, 576) нет принципиальных различий.
Что касается кровеносной системы, то полости сосудов возникли в процессе эволюции как щелевидные или каналообразные пространства в основном веществе паренхимы. Стенки кровеносных сосудов произошли путем уплотнения вокруг этих каналов основного вещества, а кровь представляет собой соединительную ткань с разжиженным основным веществом.
| 10-763 |
Итак, кольчатые черви — большая филогенетическая ветвь, поднимающаяся от низших червей. Центральную группу ветви составляет класс Polychaeta. От них в одну сторону при переходе к пресноводному и наземному образу жизни произошли малощетинковые черви, а через них и пиявки. Как это часто наблюдается, при переходе в пресные воды б развитии утрачивается метаморфоз, и оно становится прямым. Это линия в своих крайних представителях, а именно пиявках, в связи с полупаразитическим, а в некоторых случаях и паразитическим образом жизни ведет ко вторичному вытеснению целома соединительной тканью — паренхимой и к конвергенции ряда признаков с таковыми плоских червей.
Примитивные полимерные полихеты обладают неопределенно большим количеством сегментов, которые к тому же чрезвычайно сходны друг с другом (гомономная сегментация). Соответственно параподии и многие внутренние органы также многочисленны и однородны — особенность, обусловленная способом образования постларвальных сегментов во время метаморфоза личинки трохофоры. Из зоны роста формируются один за другим многочисленные сегменты, т. е. происходит множественная закладка одинаковых участков тела.
Прослеживая эволюцию отдельных филогенетических ветвей типа Annelida, мы видим характерный процесс дифференциации сегментов, параподии и внутренних метамерных органов, общее сокращение числа сегментов и органов, а часто и стабилизацию их количества. В частности, явную олигомеризацию обнаруживают перегородки целома, мета-нефридии и целомодукты, вследствие чего усиливается гетерономность сегментов путем выпадения части нефридиев и целомодуктов у многих Sedentaria. He менее обычна и частичная редукция гонад у многих по-лихет. Еще резче она выражена у Oligochaeta, где обычно имеется всего 1—2 пары мужских и близкое к этому число пар женских гонад. Не менее сильно выражена олигомеризация и у кл. Hirudinea, характеризующихся стабилизацией числа сегментов.
ТИП ЧЛЕНИСТОНОГИЕ (ARTHROPODA)
Тип членистоногих (Arthropoda) несравненно богаче всех остальных типов животных и содержит свыше 1 500 000 видов. Водные и сухопутные формы, обладающие членистыми конечностями и сегментированным телом.
Все Arthropoda характеризуются следующими основными признаками:
1. Членистоногим присуща гетерономность сегментации. Вместо гомономных, равнозначащих сегментов большинства кольчатых червей сегменты Arthropoda обладают различным строением в разных участках тела. Группы сходных сегментов выделяются в особые отделы тела, или тагмы. Чаще всего различают три тагмы: голову, грудь и брюшко. Сегменты в пределах таги, так же как и сами тагмы, могут сливаться друг с другом.
Количество сегментов, составляющих тело членистоногих, сильно варьирует в различных систематических группах. При этом проявляется тенденция к уменьшению и стабилизации числа сегментов. Наиболее постоянен сегментарный состав головного отдела (не всегда обособленного в виде самостоятельной тагмы), который состоит из головной ло-
пасти, или акрона, и четырех следующих за ней сегментов Ч Акрон гомологичен простомиуму полихет, а его придатки, антеннулы, или усики, соответствуют пальпам последних. Грудь и особенно брюшко в этом отношении более изменчивы. Заканчивается тело анальной лопастью — тельсоном, гомологичной пигидию кольчатых червей.
2. Конечности членистоногих, филогенетически развившиеся из пара-подий полихет, подвижно соединяются с телом при помощи суставов и состоят из нескольких члеников. Конечности представляют собой многоколенный рычаг, способный к сложным движениям в отличие от пара-подий кольчецов, совершающих однообразные взмахи в одной плоскости.
Конечности, расположенные на разных тагмах, зачастую специализируются для выполнения несхожих функций — захвата и измельчения пищи, движения, дыхания и т. п. Конечности брюшных сегментов у многих Arthropoda исчезают.
3. Тело членистоногих покрыто хитиновой кутикулой, образующей наружный скелет, в котором различают твердые пластинки — склериты и мягкие сочленовные мембраны. Каждый сегмент тела, как правило, покрыт 4 склеритами: дорзально располагается спинная пластинка — тергит, вентрально — брюшная пластинка, или стернит, и по бокам между ними — боковые пластинки.
Химический состав кутикулы сложен. Она включает липоиды, протеины и хитин, азотистое органическое эластичное вещество, стойкое химически. Затвердение кутикулы связано с тем, что хитин бывает пропитан углекислой известью (у ракообразных и многоножек) или инкрустирован задубленными белками (паукообразные, насекомые).
Ввиду того что тело животного одето неподатливым наружным скелетом, рост членистоногих сопровождается линьками. Старая кутикула периодически отстает от тела, а кожным эпителием выделяется новая очень мягкая кутикула; при этом старая кутикула лопается, и животное вылезает из нее, оставляя пустой чехол. В тот короткий период, пока новая кутикула мягка, и происходит увеличение размеров тела. После затвердения кутикулы, на что уходит от нескольких часов до нескольких дней, животное сохраняет приобретенные им размеры до следующей линьки.
В теле Arthropoda отсутствует мерцательный эпителий — особенность, определяемая отчасти сильной кутикуляризацией членистоногих, у которых не только покровы, но и часть кишечника, половых протоков и других органов выстлана кутикулой, исключающей возможность развития ресничек.
4. Мускулатура Arthropoda представлена отдельными мышечными пучками — мышцами, не образующими сплошного кожно-мускульного мешка. Мышцы имеют поперечнополосатую структуру.
Полость тела всех Arthropoda имеет двойственное происхождение. Во время эмбрионального развития у них в большинстве случаев закладывается сегментированный целом. Впоследствии стенки целомических мешков разрушаются, и целомические полости сливаются как друг с другом, так и с остатками первичной полости тела. Таким образом формируется смешанная полость тела, или миксоцель, в которой и располагаются внутренние органы.
1 По некоторым данным, в состав головы членистоногих входит не 4, а 5 или даже 6 сегментов (кроме акрона). Однако лишь наличие 4 задних головных сегментов не вы-зызает никаких сомнений. Они имеются у всех членистоногих и их судьба может быть прослежена во всех группах. В дальнейшем изложении мы будем учитывать только эти 4 сегмента.
10* 291
5. Пищеварительная система членистоногих состоит из трех отделов— передней, средней и задней кишок. Передний и задний отделы кишечника, будучи эктодермальными, несут кутикулярную выстилку. С разными отделами кишечного тракта связаны железы, секретирующие пищеварительные ферменты.
6. Кровеносная система членистоногих характеризуется появлением центрального пульсирующего органа — сердца, который -отсутствует у кольчатых червей. Вместе с тем кровеносная система становится незамкнутой: имеются лишь главные кровеносные сосуды — аорта и артерии, из которых гемолимфа (см. ниже) изливается в полость тела и омывает внутренние органы. Затем она вновь поступает в сосуды и сердце.
Гемолимфа — жидкость двойственной природы, она частично соответствует настоящей крови, заполняющей кровеносную систему большинства кольчатых червей, а частично — целомической жидкости. Функции гемолимфы в основном соответствуют функциям крови.
7. Органы дыхания членистоногих разнообразны. В одних случаях конечности целиком или только их части преобразуются в органы водного дыхания — жабры. Органы воздушного дыхания наземных форм — легкие — также представляют видоизмененные конечности. И, наконец, у высших членистоногих для дыхания служит особая трахейная система (с. 336).
8. Нервная система построена по типу таковой кольчатых червей и слагается из парного головного мозга, окологлоточных коннективов и брюшной нервной цепочки. Головной мозг большей частью состоит из трех отделов — протоцеребрума, дейтоцеребрума и тритоцеребрума (с. 303). Часто наблюдается концентрация ганглиев брюшной нервной цепочки и образование за счет их слияния крупных нервных узлов.
9. Выделительная система членистоногих представлена видоизмененными целомодуктами — коксальными железами или же особыми органами, возникшими в пределах типа членистоногих, а именно мальпигиевы-ми сосудами (с. 352).
10. Членистоногие обладают только половым способом размножения, причем они, как правило, раздельнополы. Нередко имеется явственный наружный половой диморфизм.
Мы примем деление членистоногих на 4 подтипа: жабродышащих (Branchiata), трахейных (Tracheata), хелицеровых (Chelicerata) и три-лобитообразных (Trilobitomorpha).
ПОДТИП ЖАБРОДЫШАЩИЕ (BRANCHIATA)
К жабродышащим относятся первичноводные членистоногие или же формы, филогенетически тесно связанные с предками, которые вели водный образ жизни. Их сегментарный состав, строение и биология разнообразны. К этому подтипу принадлежит всего один класс—Ракообразные (Crustacea).
КЛАСС I. РАКООБРАЗНЫЕ (CRUSTACEA)
Ракообразные составляют существенную часть водной фауны. Общее число известных видов не менее 20000. Ракообразные населяют главным образом моря и пресноводные водоемы различного типа, в том числе и подземные воды. Встречаются как планктонные, так и бентосные формы. Некоторые раки ведут сидячий образ жизни; известно также немало паразитов. Наконец, в разных группах раков совершался переход к наземной жизни.
От других членистоногих ракообразные отличаются некоторыми характерными особенностями. Во-первых, это одновременное присутствие двух пар усиков: придатков акрона — антеннул и видоизмененных конечностей 1-го сегмента тела — антенн '. Усики ракообразных часто обозначаются, как антенны I и антенны II соответственно. Во-вторых, только у ракообразных ножки во многих случаях сохраняют примитивное дву-ветвистое строение. В процессе эволюции, однако, этот исходный тип конечности подвергся сильным изменениям: конечности вторично становятся одноветвистыми.
Дыхание ракообразных осуществляется при помощи жабр, представляющих особые выросты ножек — эпиподиты.
Строение и физиология. Сегментация ракообразных характеризуется наибольшим среди прочих членистоногих разнообразием. Как правило, у них выделяются три отдела тела — голова, грудь и брюшко. Однако у некоторых самых примитивных форм гомономность сегментов бывает столь выражена, что грудь и брюшко почти не отличимы друг от друга (см. рис. 280, 281). Подобные исключения, правда, немногочисленны, и у большинства раков эти два отдела хорошо дифференцированы. Количество сегментов, входящих в состав тела ракообразных, подвержено сильным вариациям: от 5—8 (см. рис. 256) до 50 (см. рис. 278). В процессе эволюции ракообразных, как и других групп членистоногих, происходило уменьшение числа сегментов. Об этом говорит более богатое расчленение примитивных видов по сравнению со специализированными.
Головной отдел обладает постоянным сегментарным составом, и у всех раков включает в себя акрон с хорошо развитыми антеннулами и 4 сегмента. Первый из них — антеннальный — несет вторую пару усиков— антенны. Три же последующих сегмента снабжены видоизмененными конечностями, служащими, как правило, для захватывания и пс-ретирания пищи.
В состав груди и брюшка у различных форм входит неодинаковое число сегментов. У высших раков (подкл. Malacostraca) количество сегментов становится постоянным: в состав груди у них всегда входит 8, а в состав брюшка (за одним исключением) —6 сегментов. Следовательно, общее число их, считая с головными сегментами, равно 18 (рис.252). Заканчивается брюшко ракообразных анальной лопастью или тельсо-ном, несущим анальное отверстие. У многих примитивных форм на тель-соне часто располагаются парные придатки, образующие вилочку, или фурку (см. рис. 277).
У более примитивных форм сегменты каждого отдела тела остаются свободными и явственно отграничены друг от друга; у других же они сливаются воедино. При этом часть сегментов одного отдела может присоединяться к соседнему. В этом отношении большое разнообразие наблюдается в строении головы раков. Довольно часто сохраняется еще очень древнее расчленение головного отдела — самый передний его участок, состоящий из слившихся акрона и первого антеннального сегмента, он резко отделен от остальных сегментов. Этот участок, называемый первичной головой, или протоцефалоном, несет глаза и обе пары антенн (рис. 253, А, Б).
Противоположную картину можно наблюдать у некоторых специализированных форм (рис. 253,Б), у которых головные сегменты не только
1 У представителей подтипа Chelicerata полностью исчезают придатки акрона — антеннулы (антенны I) (с. 401); характерным признаком форм, относящихся к подтипу Tracheata (с. 331), является отсутствие у них конечностей 1-го сегмента — антенн (антенны II).

|

|
Рис. 252. Речной рак Potamobius astacus с брюшной стороны (из Резелера и Лампрехта). Л— самец; Б — самка:
I — антеннула, 2 — антенна, 3 — верхняя губа, 4— мандибула, 5 — ногочелюсть III, 6 — клешня, 7 — грудные ходные ноги, 8 — мужское половое отверстие, 9 — совокупительные ножки, 10 — брюшные ножки, // — плавательная хвостовая ножка, 12 — анальное отверстие, 13 — отверстие антеннальной железы, 14 — максиллы II, 15 — ногочелюсть II, 16 — женское половое отверстие, 17 — рудиментарная ножка

|
| В |
Рис. 253. Расчленение головы ракообразных (по Снодграссу). А — жаброног Eubranchipus vernatis (подкл. Branchiopoda), протоцефалон обособлен; Б — креветка Spirontocaris polaris (отр. Decapoda), протоцефалон отпрепарирован; В — бокоплав Orchestoidea calif arnica (отр. Amphipoda), голова сложная:
1 — антеннула, 2 — антенна, 3 —верхняя губа, 4 — сложный глаз, 5^ мандибула, 6 — грудные ножки, 7 — протоцефалон, 8 — 2-й сегмент головы, 9 — 3-й и 4-й сегменты головы. 10 —1-й грудной сегмент, // — 2-й грудной сегмент, 12 — максиллы I, 13 — максиллы II, 14 — ногочелюсть, принадлежащая первому грудному сегменту, вошедшему в состав головы
сливаются друг с другом, но к ним присоединяются еще 1 или 2 грудных сегмента. Все вместе они образуют единое компактное целое.
Голова ракообразных несет ряд придатков, имеющих различное происхождение, к которым иногда присоединяются также особые непарные выросты ее стенок.

|
Ротовое отверстие спереди прикрывается особой непарной кутику-лярной складкой — верхней губой (рис. 253, А, В). Очень часто спинной и боковые края заднего сегмента головы сильно выдаются в виде плоских выростов, образуя так называемый головной щит, или карапакс. Во многих случаях карапакс сильно разрастается назад и может более или менее полно закрывать со спинной стороны и с боков сегменты туловища, а иногда образует даже двустворчатую раковину, в которой помещается все тело (см. рис. 256, 279). У высших раков подкласса Malacostraca карапакс срастается с сегментами груди.
| Рис. 254. Схема строения примитивной конечности ракообразных (по Снодграссу): 1 — протоподит, 2 — эн-доподит, 3 — экзоподит, 4 — эпиподиты — дыхательные придатки |
Парные придатки головы представлены антен-нулами и четырьмя парами видоизмененных конечностей. Антеннулы, или антенны I, принадлежат головной лопасти — акрону — и гомологичны паль-пам Polychaeta. Они иннервируются от средней части мозга — дейтоцеребрума и расположены на передней стороне головы, впереди ротового отверстия. Антеннулы обычно одноветвисты. Лишь у некоторых высших раков они вторично расщепляются на две (например, у речного рака) или даже на три ветви. Чаще всего антеннулы функционируют как
органы осязания и обоняния, хотя иногда служат и для плавания (см. рис. 282).
На следующих за акроном сегментах располагаются уже настоящие конечности, гомологичные параподиям кольчецов. Из всех членистоногих у раков сохраняется наиболее примитивный тип строения ножек, которые во многих случаях двуветвистые (рис. 254), чем они напоминают параподии полихет. Такая конечность состоит из расчлененного основания—протоподита, от конца которого отходят 2 ветви: внутренняя, более близкая к медианной линии тела — эндоподит, и наружная — экзоподит. Кроме того, протоподит несет особые выросты — эпиподиты, выполняющие функцию органов дыхания — жабр (рис. 254).
Этот исходный тип конечности претерпевает, однако, значительные изменения, связанные, в первую очередь, с дифференциацией функций: питание, передвижение и т. п. Последнее обстоятельство является причиной того, что конечности, расположенные на разных отделах тела, иногда довольно сильно отличаются друг от друга.
Антенны вторые, или просто антенны, представляют конечности первого головного сегмента. Они иннервируются от тритоцеребральной части мозга, а у очень примитивных раков — даже от окологлоточных коннективов. Антенны играют различную роль. У водяных блох (рис. 255), относящихся к листоногим ракам (отр. Phyllopoda), они служат для плавания. У большинства высших раков — это органы чувств. Так, у речного рака эндоподит антенны образует длинный членистый чувствительный «бич», тогда как экзоподит имеет вид короткой защитной пластинки (см. рис. 252).
За антеннами II следуют три пары конечностей остальных сегментов головы. Вторая пара головных конечностей, жвалы или мандибулы, иг-
рает главную роль в размельчении пищи. У характерной личинки многих ракообразных — науплиуса (с. 309) — это типичная двуветвистая конечность, основание которой обладает, однако, особым жевательным отростком. Во взрослом состоянии такая форма мандибул сохраняется редко. Обычно обе ветви редуцируются (или эндоподит сохраняется в виде короткого щупика), а протоподит вместе с его жевательным отростком образует толстую зазубренную верхнюю челюсть, к которой прикрепляются мощные мышцы. За мандибулами располагаются конечности третьего и четвертого головных сегментов — две пары нижних челюстей, или максиллы первые и максиллы вторые. Они чаще всего имеют вид нежных листообразных ножек с несколько редуцированными ветвями и с жевательными отростками на члениках протоподита.
Грудные конечности раков очень разнообразны и зачастую выполняют различные функции. Во многих случаях, однако, они являются органами движения (плавание, передвижение по твердому субстрату). Часто конечности бывают двуветвисты, но нередко экзоподит редуцируется (например, у речного рака; см. рис. 252). У большого числа форм ножки

|
одного — трех передних грудных сегментов принимают участие в поддерживании пищи и ее размельчении. В этом случае они становятся более или менее похожими на нижние челюсти и соответственно называются ногочс-люстями.
| 13- |
Брюшные конечности имеются только у высших раков (подкл. Malacostraca) и большей частью двуветвисты. Эти придатки нередко несут не двигательную, а какую-нибудь иную функцию: дыхательную, органов совокупления и т. д. У многих десятиногих раков (отр. Decapoda) последняя пара брюшных двуветвистых конечностей превратилась в мощные пластинчатые хвостовые ноги (см. рис. 252), играющие вместе с уплощенным тельсоном важную роль при плавании задом наперед. У некоторых паразитических ракообразных конечности туловища сильно редуцируются вплоть до полного исчезновения (Sacculina, самки Dendrogaster; см. рис. 289; 290,£ и др.).
| Рис. 255. Самка водяной блохи Daphnia ри-lex (из Лилиенборга): / — сложный глаз, 2 — науплиальяый глаз, 3 — мозг, 4 — брюшной край головы. 5 — антеннула, 6 — антенна, 7 — верхняя челюсть (мандибула), 8 — кишка, 9 — печеночный вырост, 10 — ыаксил-лярная выделительная железо* it — сердце. 12 — яичник, 13 — выводковая камера. 14 — спинные выросты брюшка, закрывающие выход в выводковую камеру. 15 — задний вырост. 16 — щетинки, 17 — брюшко, 18 — грудные ножки, 19 — 1-я пара грудных ножек |
Покровы ракообразных состоят из кутикулы и подлежащих слоев: гиподермального эпителия (гиподермы) и базальной мембраны. Кутикула, выделяемая клетками гиподермы, представляет собой сложное образование, состоящее из нескольких слоев. В периферических слоях кутику-
лы откладывается известь, в результате чего покровы у многих видов становятся более жесткими и прочными. Внутренний слой состоит в основном из мягкого и эластичного хитина. У Crustacea в отличие от паукообразных и насекомых отсутствует самый наружный слой кутикулы, препятствующий испарению воды из организма (с. 350), что связано с первичноводным образом их жизни. Эта особенность сохраняется и у тех форм, которые перешли к жизни на суше (мокрицы, см. рис. 295.Л; некоторые сухопутные крабы и т. п.). Поэтому обитают они только в тех местах, где влажность воздуха близка к полному насыщению.
Кутикула выполняет функции наружного скелета. У мелких низших форм скелет этот большей частью остается довольно мягким и прозрачным, у высших нередко кутикула становится более толстой и превращается в твердый панцирь. Скелет имеет двоякое значение: во-первых, он защищает животное от внешних воздействий, и, во-вторых, дает точки опоры для прикрепления различных мышц. Местами на внутренней поверхности скелета образуются отростки в виде гребней и перекладин, которые служат специально для прикрепления мускульных пучков.
Мускулатура ракообразных, как и у всех членистоногих, состоит из поперечнополосатых волокон. Она не образует сплошного кожно-мус-кульного мешка, но распадается на отдельные более или менее мощные мышечные пучки, соединяющие между собой различные точки внутренней поверхности скелета. Мышцы располагаются чаще всего так, что один конец мускула прикрепляется к стенке одного сегмента тела или членика конечности, другой — к стенке другого. У раков, обладающих двустворчатым панцирем (подкл. ракушковые раки — Ostracoda; рис. 256), имеется особая замыкательная мышца, идущая от одной створки к другой, поперек тела. Полость тела представлена миксоцелем.

|
| 12 |
| 11 |
Рис. 256. Ракушковый рачок Heterocypris reptans (по Волтереку):
Глаз, 2— антеннула, 3—антенна, 4 — пищевод, 5 — мандибула, 6 — мак-силла, 7—1-я грудная конечность, 8 — запирательный мускул, 9 — 2-я грудная конечность, 10 — печеночный вырост кишки, 11 — вилочка, 12 — яичник, 13 — зачатковая зона яичника, 14 — 3-я грудная конечность, 15 — яйцевод, наполненный яйцами, 16 — средняя кишка
Пищеварительная система развита хорошо (рис. 257). Кишечный канал имеет вид прямой или слегка согнутой трубки и состоит из передней, средней и задней кишок. Порошица открывается на брюшной стороне

|
анальной лопасти (тельсона). Передняя и задняя кишки выстланы продолжением общей кутикулы, покрывающей тело снаружи. Во время линьки выстилка этих частей кишечника тоже линяет, выходя изо рта и порошицы в виде двух кутику-лярных трубок. Кутикула, выстилающая стенки передней кишки, может образовывать местные утолщения, служащие для перетирания пищи. У некоторых форм эти утолщения имеют вид волосков и шипиков, но особенно сильного развития они достигают у высших раков, относящихся к отр. Decapoda. Так, у речного рака передняя кишка в своей конечной части образует большое расширение — желудок, который подразделяется на два отдела — кардиальный, или жевательный, и пилорический (рис. 258). На спинной и боковых стенках кардиальной части желудка кутикула утолщается в три мощные, пропитанные известью жевательные пластинки, сильно зазубренные по свободному краю. В пилорической части желудка тонкие кутикулярные выросты образуют подобие фильтра, через который проходит только сильно измельченная пища.
| Рис. 257. Пищеварительный канал речного рака (по Гексли): / — желудок, 2 — пилориче-ская часть желудка, 3 — слепой дорзальный вырост средней кишки, 4 — средняя кишка, 5 — валик, отделяющий среднюю кишку от задней, 6 — задняя кишка, 7 — проток печени, 8 и 9 — гребни на стенке желудка |
Средняя кишка иногда (отр. Copepoda) имеет вид простой трубки, но чаще образует боковые выпячивания, обладающие железистой природой. В этих выростах не только выделяются пищеварительные ферменты, но и переваривается жидкая пищевая кашица.
Большей частью бывает пара таких выростов, причем каждый из них может более или менее сильно ветвиться. Это так называемые печеночные придатки, образующие в случае сильного развития массивную печень. В зачаточном состоянии они имеются у некоторых водяных блох Gladocera (см. рис. 255). У бокоплавов
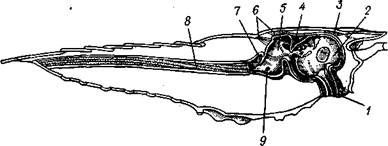
Рис. 258. Продольный разрез речного рака, показывающий расположение основных частей пищеварительной системы (по Гексли):
I — пищевод, 2 — кардиальная часть желудка, 3 — передние мышцы желудка,
4 — срединный зуб желудка, 5 — пилорическая часть желудка, 6 — задние
мышцы желудка, 7 — слепой дорзальный вырост средней кишки, S — задняя
кишка, 9 — отверстие протока печени
и равноногих раков (отр. Amphipoda и отр. Isopoda) они имеют характер двух пар длинных трубчатых мешков (рис. 259). У речного рака крупная двухлопастная печень состоит из множества мелких трубочек, собирающихся с каждой стороны в один проток, впадающий в среднюю кишку. Секрет печени у речного рака поступает затем из средней кишки в жевательный желудок. Физиологические исследования секрета «печени» раков показывают, что название ее не совсем правильно; функционально ее можно сравнивать только с совокупностью печени и поджелудочной железы позвоночных. Секрет печени рака может не только подобно желчи расщеплять жиры, переводя их в состояние эмульсии, но и расщепляет белки и углеводы. Кроме секреторной функции печень раков способна к фагоцитозу, ее клетки захватывают мелкие частицы пищи и переваривают их внутри-клеточно. Часто можно наблюдать известную корреляцию между размерами печеночных придатков и длиной средней кишки. Чем слабее печеночные придатки (например, у водяных блох), тем длиннее средняя кишка, и наоборот, например, у речного рака средняя кишка составляет 1/20 часть всей длины кишечника.
Задняя кишка образует прямую трубку, лишенную всяких придатков.
У некоторых паразитических раков (Sacculina и др.) кишечник полностью атрофирован.
Пищей ракам служат чаще всего различные более мелкие животные или же разлагающиеся животные и растительные остатки.

|
| Рис. 259. Изолированная пищеварительная система водяного ослика Asellus aquaticus (из Сарса): 7 — жевательный желудок, 2 — средняя кишка, 3 — печеночные выросты. 4 — задняя кишка |
Дыхательная система ракообразных, как и у класса Polychaeta, находится обыкновенно в тесной связи с конечностями. У многих мелких ракообразных специальные органы дыхания отсутствуют (подкл. Ostracoda, Maxillopoda и др.)—дыхание совершается всей поверхностью тела. У других раков для дыхания служат кожные жабры. Это эпиподиты, пластинчатые или ветвистые тонкостенные выросты, отходящие от протоподитов ног. Чаще всего они развиты на грудных конечностях. У представителей отряда Decapoda жабры сидят под боковыми краями головного щита внутри особой жаберной полости; развиваясь сначала на про-топодитах грудных ног, они впоследствии частью переходят и на боковую стенку самого тела (рис. 260). При этом жабры Decapoda располагаются продольными рядами. Жабры одного ряда сохраняют первичное положение на протоподитах ног, жабры другого помещаются на местах соединения протоподитов с телом, тогда как в третьем ряду они сидят уже на боковой стенке тела. Вода входит в жаберные полости с одного конца через щель между головным щитом и телом, а выталкивается из них с другого, причем направление тока воды (спереди назад или обратно) может меняться. Проведению воды через жаберные полости способствует движение особого отростка максилл вторых, который производит до 200 машущих движений в 1 мин. В жабрах продолжается полость тела, в которую попадает гемолимфа; кутикула жабр очень нежна, и через нее легко происходит газообмен.
Интересные изменения дыхательного аппарата
|
наблюдаются у некоторых раков, перешедших к наземному образу жизни. У части наземных Isopoda, а именно у мокриц (см. рис. 295,Л), на пластинчатых брюшных ногах имеются глубокие ветвящиеся впячивания покровов. Полость конечности заполнена гемолимфой, омывающей впячивания. Внутрь впячиваний входит воздух и диффундирует в окружающую гемолимфу. Эти органы сильно напоминают трахеи, т. е. органы дыхания типичных сухопутных членистоногих (с. 336).
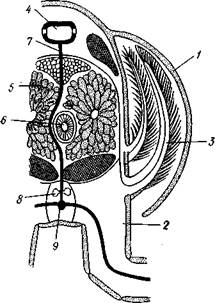
Кровеносная система ракообразных (рис. 261), как и прочих членистоногих, не замкнута: частично ге-молимфа движется внутри сосудов, выстланных собственным эпителием, частично же в участках полости тела, не ограниченных специальными стенками, — синусах.
Кровеносная система находится

Рис. 260. Схема поперечного разреза через челюстегрудь речного рака в области сердца (из Матвеева):
Жаберная крышка — боковой край кара-пакса, 2 — основание ноги, 3 — жабра, 4 —сердце, 5 — печень, 6 — кишка, 7 — нисходящая артерия, 8 — нервная цепочка, 9 — поднервная продольная артерия
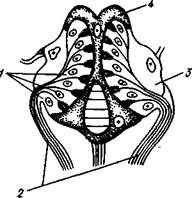
Рис. 261. Схема кровеносной системы
Речного рака (по Гегенбауэру): / — антеннальная, сяжковая артерия, 2 — передняя аорта (глазная артерия), 3 — сердце, 4 — перикардий, 5 — жаберносердечные каналы, 6 — нисходящая артерия, 7 — задняя (верхняя брюшная) артерия, 8 — поднервная артерия, 9 — брюшной венозный синус
в известной зависимости от степени развития органов дыхания. Обыкновенно кровеносная система довольно сложна, но там, где дыхание совершается через всю поверхность тела, от нее сохраняется только сердце (см. рис. 255) или же кровеносная система исчезает целиком.
У более примитивных форм сердце имеет метамерное строение: оно образует трубку, идущую по спинной стороне вдоль всего тела (у не-которых жаброногих раков) и снабженную в каждом сегменте парой остий. Однако у водяных блох (из отр. Phyllopoda) наблюдается укорачивание сердца до степени бочонковидного мешочка всего с одной парой остий, служащего лишь для перемешивания гемолимфы. Среди высших раков также имеются формы как с длинным трубчатым сердцем (отр. Amphipoda и особенно ротоногие раки — отр. Stomatopoda), так и с укороченным. У представителей отр. Decapoda это небольшой мешочек с тремя парами остий.
Зависимость кровеносной системы от дыхательной ярче всего сказывается в относительном положении сердца и жаберных придатков. В тех случаях, когда жабрами служат эпиподиты грудных ног, сердце целиком залегает в груди, когда же для дыхания приспособлены эпиподиты брюшных конечностей, сердце располагается в брюшке.
От переднего, а нередко и от заднего конца сердца отходит по крупному сосуду — передняя и задняя аорты. Детали кровообращения сильно варьируют. Примером хорошо развитой кровеносной системы может служить таковая речного рака (рис. 261), у которого от сердца, лежащего в перикардии, отходят несколько крупных сосудов: передняя аорта, антеннальные, или сяжковые, артерии, верхняя брюшная и нисходящая артерии и т. д. Сосуды, отходящие от сердца, сначала ветвятся, потом обрываются, так что гемолимфа изливается прямо в полость тела и там постепенно отдает кислород. Из полости тела гемолимфа по хорошо развитой системе венозных синусов течет в жабры, где и обогащается кислородом. Оттуда по особым жаберно-сердечным каналам она направляется в перикардий. Последний, как и у других Arthropoda, представляет собой обособленный участок полости тела, окружающий сердце. У речного рака перикардий замкнут, и в него вливаются лишь жаберно-сердечные сосуды, у других же представителей ракообразных он широко соединяется с остальной полостью тела. Из перикардия гемолимфа через остии поступает в сердце. Гемолимфа ракообразных во многих случаях бесцветна, но у многих окрашена в красный цвет гемоглобином, растворенным в плазме. У некоторых десятиногих раков (часть крабов) гемолимфа синеватая или же синеет при соприкосновении с воздухом: это связано с присутствием в гемолимфе дыхательного пигмента — гемоцианина, осуществляющего транспорт кислорода и содержащего медь.

|
Нервная система раков напоминает таковую кольчатых червей. Она состоит из парного головного мозга, окологлоточных коннективов и пары брюшных нервных стволов с ганглиями в каждом сегменте. Именно такова очень примитивная нервная система многих представителей подкл. жаброногих раков — Branchiopoda (рис. 262,Л), у них оба брюшных ствола еще далеко отставлены друг от друга, как у многих полихет.
| Рис. 262. Типы строения центрального нервного аппарата ракообразных (по Гизбрехту). Л — отр. Anostraca; Б -отр. Euphausiacea; В - отр. Sto- |
У большинства раков из
менение нервной системы
происходило в двух направ
лениях. Во-первых, брюш
ные нервные стволы сблизи
лись, правый и левый ганг
лий каждого сегмента сли
лись, вследствие чего попе
речные комиссуры между
ними исчезли, и только
двойственность продольных
перемычек между ганглия
ми соседних сегментов на
поминает о парном проис
хождении брЮШНОГО нерв-
НОГО СТВОЛа. Образовалась
так называемая брюшная
нервная цепочка (рис.
262,5). Второе изменение— Copepoda; Ж — подкл. Ostracoda
продольная концентрация нервной цепочки. По мере наступающего у разных групп раков тесного слияния отдельных сегментов тела друг с другом произошло сближение ганглиев в продольном направлении, укорачивание продольных стволов и слияние воедино нервных узлов, принадлежащих разным сегментам (рис. 262, Б, В). Так, у речного рака, несмотря на состав тела из 18 сегментов, вне головного мозга, иннервирующего антеннулы и антенны, т. е. в составе брюшной нервной цепочки, остается всего 12 нервных узлов, а именно: подглоточный узел (продукт слияния ганглиев, отвечающих трем парам ротовых конечностей и трем парам ногочелюстей), 5 грудных узлов (отвечающих такому же числу сегментов ходных ног) и 6 брюшных ганглиев, причем последний из них иннервирует не только свой сегмент, но и тельсон (рис. 262.Г). Очень далеко заходит концентрация нервной системы у крабов (рис. 262.Д); у них имеются лишь две нервные массы — головной мозг и общая масса в груди, образовавшаяся в результате слияния всех нервных узлов брюшной цепочки. Значительная степень концентрации, нервной системы свойственна также веслоногим ракам (отр. Copepoda) и ракушковым ракам (подкл. Ostracoda), у которых образуется ком-

|

|
| Хроматофоры |
| Сахар крови. |
Рис. 263. Схема нейросекретор-
ной системы синусовой железы
краба (из Велша):
/ — брюшная ганглнозная масса, 2 — головной мозг, 3 — зрительный нерв, 4 — глазной стебелек, 5 — ней-росекреторные клетки, в — отростки нейросекреторных клеток, 7 — сложный глаз, 8 — синусовая железа, 9 — эндокринная железа
Рис. 264. Продольный разрез через антеннулу глубоководного краба Geryon affinis (из Дофлейна).
Каждый волосок получает нервное волокно от нервной клетки у своего основания
пактная ганглиозная масса, пронизанная каналом пищеварительной системы (рис. 262, Е, Ж).
Строение головного мозга раков довольно сложно: он состоит из парных долей — протоцеребрума и дейтоцеребрума. Обычно с головным мозгом сливаются и смещающиеся вперед ганглии сегмента антенн II. В этом случае образуется третий отдел мозга — тритоцеребрум. От протоцеребрума к глазам направляются зрительные нервы. По ходу последних выделяются ганглиозные скопления нервных клеток. Антеннулы иннервируются от дейтоцеребрума. Нервы к антеннам II чаще всего отходят от окологлоточных коннективов. У раков имеется хорошо развитая симпатическая нервная система, главным образом иннервирую-щая кишечник. Она состоит из церебрального отдела и непарного симпатического нерва. По ходу последнего располагается несколько ганглиев.
У ракообразных, как и у других членистоногих, в состав нервной системы входят специальные клетки, способные выделять особые гормоны — нейросекреты. Последние поступают в гемолимфу и влияют на деятельность отдельных органов, обмен веществ, процессы линьки, метаморфоза и т. д. Нейросекреторные клетки раков расположены в различных частях протоцеребрума, тритоцеребрума и ганглиев брюшной нервной цепочки. Важную роль играют нейросекреторные клетки, образующие группы по ходу зрительного нерва (рис. 263). Секрет этих клеток по отросткам поступает в особое образование — синусовую железу, а оттуда в гемолимфу.
| Рис. 265. Антеннула речного рака (из Шимкевича): 1 к 2 — два жгута усика. 3 — основные членики, 4 — отверстие ямки статоциста, 5 — чувствительные волоски на дне статоциста, 6 — песчинки внутри статоциста, 7 — нерв. Оба жгута усажены осязательными и обонятельными волосками |
Органы чувств хорошо развиты. Чувство осязания приурочено только к определенным точкам покровов, а именно к части волосков и щетинок на поверхности антеннул, антенн и других конечностей. У основания таких чувствительных волосков под гиподермальным эпителием лежат биполярные нервные клетки. Несколько измененные, отличающиеся особенно проницаемой кутикулой волоски на антеннулах многих раков являются органами химического чувства (рис. 264). Органы равновесия встречаются у ракообразных сравнительно редко. Однако у большинства десятиногих раков в основном членике антеннул имеется статоцист — глубокое впячивание покровов, усаженное внутри нежными перистыми чувствительными волосками (рис. 265). Чаще всего впячивание непосредственно сообщается с внешней средой, в него легко попадают мел-

кие песчинки, играющие роль «слуховых камешков», или «статолитов». При линьке хитиновая выстилка и статолиты открытых впячиваний удаляются, и животное набирает новый запас песчинок либо при помощи клешней, либо засовывая голову многократно в песок. Кроме декапод
|
статоцистами обладают и некоторые другие высшие раки.
Особенно интересны у ракообразных глаза, встречающиеся в двух формах. Во-первых, нередко имеется непарный лобный глаз, называемый часто науплиаль-ным' глазом потому, что он характерен для личинки науплиуса, хотя встречается и у взрослых раков. Науплиальный глаз лежит между основаниями антеннул и представляет собой продукт слияния 2 или даже 4 глазных бокальчиков, состоящих из одного слоя ретинальных клеток (рис. 266, А). Каждый бокальчик окружен как бы шапочкой темного пигмента. Выемку бокала, обращенную к поверхно-

Рис. 266. Органы зрения ракообразных. А — схема строения непарного глазка Cypris (из Бючли); Б — разрез через сложный глаз жабронога
Branchipus (из Клауса); В — участок разреза через сложный глаз: / — ретинальные клетки, 2 — зрительный нерв, 3—хрусталик, 4 — пигментные клетки, 5 — хрустальные конусы, 6 — слой ретинальных и пигментных клеток, 7 — нервные волокна, 8 — мускул глазного стебелька, 9 — мозг. 10, II — ганглиоз-ные скопления нервных клеток, 12 — хитиновая прозрачная кутикула, 13 — ди-стальные части ретинальных клеток, 14 — ретинальные клетки, 15 — базальная мембрана, 16 — нервные волокна, отходящие от базальной части ретинальных клеток, 17 — прослойки пигмента между отдельными омматидиями
сти тела, занимает светопреломляющий хрусталик. Глаз имеет инвертированное строение, так как нервные волокна отходят от концов ретинальных клеток, обращенных в полость глазного бокала.
Во-вторых, большинство раков обладает парой сложных, или фасе-тированных, глаз (рис. 266,5), очень характерных для большинства членистоногих. Каждый такой глаз содержит множество мелких глазков, или омматидиев (у речного рака их больше 3000), тесно сближенных и отделенных друг от друга лишь тонкими прослойками пигмента.
Омматидий (рис. 266.В), в свою очередь, состоит из сложно устроенного светопреломляющего аппарата, группы чувствительных ретинальных клеток, от которых отходят нервные окончания, дающие начало зрительному нерву, и экранирующих пигментных клеток'.
Сложные глаза часто сидят на особых подвижных выростах головы, стебельках. У различных представителей раков могут быть во взрослом состоянии следующие комбинации глаз: 1) только науплиальный глаз

|

|
| * * |
Рис. 267. Действие гормона синусовой железы на хроматофоры креветки Le-ander. A — пигментные клетки; Б — посветлевшие под влиянием гормонов пигментные клетки (из Ганшт-рема)
Рис. 268. Антеннальная железа речного рака (из Вальса):

