




размельчаетс€ и разминаетс€ пища, тогда как прочие членики составл€≠ют род щупальца. ” представителей некоторых отр€дов (скорпионы, лжескорпионы) педипальпы превращены в мощные длинные клешни (см. рис. 389, 390,∆), у других Ч похожи на ходные ноги (см. рис. 390, ≈, ƒ; рис. 393).
ќстальные 4 пары головогрудных конечностей состо€т из 6Ч7 чле≠ников и играют роль ходных ног. ќни заканчиваютс€ коготками.

|
| –ис. 391. —амец и самка каракур≠та Latrodectus tredecimguttatus (по ћариковскому) |
” взрослых паукообразных брюшко лишено типичных конечностей, хот€ они, несомненно, произошли от пред≠ков, обладавших хорошо развитыми ножками на передних брюшных сег≠ментах. ” эмбрионов многих паукооб≠разных (скорпионов, пауков) на брюш≠ке закладываютс€ зачатки ножек, ко≠торые лишь впоследствии подвергают≠с€ регрессу. ¬прочем, и во взрослом состо€нии брюшные ножки иногда со≠хран€ютс€, но в модифицированном виде. “ак, у скорпионов на первом сег≠менте брюшка есть пара половых кры≠шечек, под которыми открываетс€ по≠ловое отверстие, на втором Ч пара гребенчатых органов (см. рис. 389), ко≠торые снабжены многочисленными нервными окончани€ми и играют роль ос€зательных придатков. » те и другие представл€ют видоизмененные конеч≠ности. “акова же природа и легочных мешков, расположенных на сегментах брюшка у скорпионов, некоторых пау≠ков и лжескорпионов (с. 413), ќт ко≠нечностей берут свое начало и паутин≠ные бородавки пауков. Ќа нижней по≠верхности брюшка впереди порошицы у них находитс€ 2Ч3 пары бугорков, усаженных волосками и несущих труб-ковидные протоки многочисленных пау≠тинных желез (рис. 394). √омологи€ этих паутинных бородавок брюшным конечност€м доказываетс€ не только их эмбриональным развитием, но и строением их у некоторых тропических пауков, у которых бородавки особенно сильно развиты, состо€т из нескольких члеников и даже по виду напо≠минают ножки.
ѕокровы хелицеровых состо€т из кутикулы и подлежащих слоев: гиподермального эпители€ (гиподермы) и базальной мембраны. —ама кутикула представл€ет сложное трехслойное образование. —наружи рас≠полагаетс€ липопротеиновый слой, надежно предохран€ющий организм от потери влаги при испарении. Ёто позволило хелицеровым стать на≠сто€щей сухопутной группой и заселить самые засушливые районы зем≠ного шара. ѕрочность кутикуле придают белки, задубленные фенолами и инкрустирующие хитин.

|

|
ѕроизводными кожного эпите≠ли€ €вл€ютс€ некоторые желези≠стые образовани€, в том числе €довитые и паутинные железы. ѕервые свойственны паукам, жгу-

–ис. 392. —обачий клещ Ixodes п-cinus. A Ч самка, вид со спинной стороны (по ѕомеранцеву); Ѕ Ч ротовые органы со спинной сторо≠ны; ¬ Ч то же, с брюшной сторо≠ны (из ƒогел€):
/ Ч хоботок, 2 Ч туловище, 3 Ч ходные ноги, 4 Ч основание хоботка, 5 Ч педи-пальпа, 6 Ч гипостом, вырост основа≠ни€ хоботка, 7 Ч хслицеры. 8 Ч крою≠щие пластинки
–ис. 393. –отовые органы пау≠ка крестовика Araneus diade-matus (по »ванову):
1 Ч конечный когтевидный членик хелицеры, 2 Ч основной членик хе-лицеры, 3 Ч педнпальпа, 4 Ч жева≠тельный вырост основного членика педипальпы, 5 Ч основной членик ходной ноги
|
|
|
|
| 13 |
| 8 |
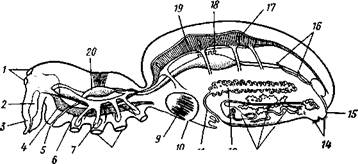
–ис. 394. —хема организации паука (отр. Aranei) (из јверинцева): I Ч глаза, 2 Ч €довита€ железа, 3 Ч хелицера, 4 Ч мозг, 5 Ч рот, 6 Ч под-глоточный нервный узел, 7 Ч железистый вырост кишечника, S Ч основа≠ни€ ходных ног, 9 Ч легкое, 10 Ч легочное отверстие Ч дыхальце, II Ч €й≠цевод, 12 Ч €ичник, 13 Ч паутинные железы, 14 Ч паутинные бородавки, 15 Ч анус, 16 Ч мальпигиевы сосуды. /7 Ч остии, 18 Ч протоки печени, 19 Ч сердце, 10 Ч глотка, св€занна€ со стенкой тела мускулатурой
топогим и скорпионам; вторые Ч паукам, лжескорпионам и некоторым клещам.

|
ѕищеварительна€ система у представителей разных отр€дов хелице-ровых сильно варьирует. ѕередн€€ кишка обыкновенно образует расши≠рениеЧ снабженную сильными мышцами глотку, котора€ служит в ка≠честве насоса, вт€гивающего полужидкую пищу, так как паукообразные не принимают твердую пищу кусками. ¬ переднюю кишку открываетс€ пара небольших Ђслюнных железї. ” пауков секрет этих желез и печени способен энергично расщепл€ть белки. ќн вводитс€ в тело убитой до≠бычи и приводит содержимое ее в состо€ние жидкой кашипы, котора€ затем всасываетс€ пауком. «десь имеет место так на≠зываемое внекишечное пищеварение.
| II |

|
” большинства паукообразных средн€€ кишка об≠разует длинные боковые вып€чивани€, увеличиваю≠щие вместимость и всасывающую поверхность кишеч≠ника. “ак, у пауков (рис. 394) от головогрудной части средней кишки к основани€м конечностей идут 5 пар слепых железистых мешков; аналогичные вып€чива≠ни€ имеютс€ у клещей, сенокосцев и других паукооб≠разных. ¬ брюшной отдел средней кишки открывают≠с€ протоки парной пищеварительной железы Ч печени (рис. 395); она выдел€ет пищеварительные ферменты и служит дл€ всасывани€ питательных веществ. ¬ клетках печени происходит внутриклеточное пище≠варение.
ќгромное большинство Arachnida Ч хищники, но имеютс€ паразитические формы, питающиес€ кровью позвоночных, и много растительно€дных форм, сосу≠щих соки растений или поедающих растительные остатки.
¬ыделительна€ система паукообразных по сравне≠нию с мечехвостами имеет совершенно иной характер. Ќа границе между средней и задней кишкой в пищева≠рительный канал открываетс€ пара большей частью ветв€щихс€ мальпигиевых сосудов. ¬ отличие от “га-cheata они энтодермального происхождени€, т. е. обра≠зуютс€ за счет средней кишки (см. рис. 394, 395). ак в клетках, так и в просвете мальпигиевых сосудов есть многочисленные зерна гуанина Ч главного продукта выделени€ паукообразных. √уанин, как и мочева€ кис≠лота, экскретируема€ насекомыми (с. 352), обладает малой растворимостью и удал€етс€ из организма в ви≠де кристаллов. ѕотери влаги при этом минимальные (с. 353), что важно дл€ животных, перешедших к жиз≠ни на суше.
| –ис. 395. ѕищева≠ рительна€ система скорпиона (по ѕавловскому): / Ч средн€€ кншка, 2 Ч печень. я Ч про≠токи печени, 4 Ч дор-зо-вентральные мыш≠цы, пронизывающие печень, S Ч мальпн-гиев сосуд, в Ч зад≠н€€ кишка |
роме мальпигиевых сосудов паукообразные обла≠дают еще и типичными коксальными железами Ч пар≠ными мешковидными образовани€ми мезодермальной природы, лежащими в двух (реже в одном) сегментах головогруди. ќни хорошо развиты у зародышей и в молодом возрасте, но у взрослых животных более или менее атрофируютс€. ¬полне сформированные кок-сальные железы (рис. 396) состо€т из концевого эпи≠телиального мешочка, петлевидно извитого канала и более пр€мого выводного протока с мочевым пузырь-
|
|
|
ком и наружным отверстием. онцевой мешочек отвечает ресничной во≠ронке целомодукта, отверстие которой замкнуто остатком целомическо-го эпители€. оксальные железы открываютс€ у основани€ 3-й или 5-й пары конечностей.
|
Ќервна€ система Arachnida разнообразна. Ѕудучи св€занной в про≠исхождении с брюшной нервной цепочкой кольчатых червей, у паукооб≠разных она обнаруживает четко выраженную тенденцию к кон≠центрации.
√оловной мозг имеет сложное строение. ¬ его состав вход€т два отдела: передний, иннервирую-щий глаза, Ч протоцеребрум и задний Ч тритоцеребрум, посыла≠ющий нервы к первой паре конеч-

–ис. 396. оксальные железы сенокос≠ца Garella variegata (из Ћанга):
/ Ч основани€ ходных ног, 2 Ч трахейный ствол, 3 Ч выделительное отверстие, 4 Ч перерезанный выводной канал железы (на левой стороне он сохранилс€ полностью), 5 Ч стигма, 6 Ч резервуар железы (моче≠вой пузырек)
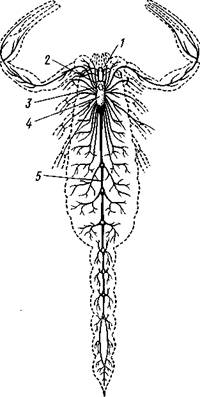
–ис. 397. Ќервна€ система скорпиона Androctonus (из √анштрома):
/ Ч глаза, 2 Ч мозг, 3Ч окологлоточные
коннективы, 4 Ч подглоточна€ ганглиозна€
масса, 5 Ч брюшна€ нервна€ цепочка
ностей Ч хелицерам. ’арактерный дл€ других членистоногих (ракооб≠разные, насекомые) промежуточный отдел мозга Ч дейтоцеребрум у паукообразных отсутствует. —в€зано это с исчезновением у них, как и у остальных хелицеровых, придатков акрона Ч антеннул, или усиков, которые иннервируютс€ именно от дейтоцеребрума.
ћетамерность брюшной нервной цепочки всего €снее сохран€етс€ у скорпионов (рис. 397). ” них имеетс€ помимо головного мозга и около≠глоточных коннективов больша€ ганглиозна€ масса в головогруди на брюшной стороне, дающа€ нервы к 2Ч6-й парам конечносте'й и 7 гангли≠ев, на прот€жении брюшного отдела нервной цепочки. ” сольпуг кроме сложного головогрудного гангли€ на нервной цепочке сохран€етс€ е:ле один узел, а у пауков уже вс€ цепочка слилась в головогрудный ганглий.
Ќаконец, у сенокосцев и клещей нет даже €вственного разграничени€ между головным мозгом и головогрудным ганглием (рис. 398), так что нервна€ система образует вокруг пищевода сплошное ганглиозное кольцо.
|
ќрганы чувств Arachnida разнообразны. ќчень важные дл€ пауко≠образных механические, ос€зательные раздражени€ воспринимаютс€ различно устроенными чувствительными во≠лосками, которые особенно многочисленны на педипальпах. —пециальные волоски Ч трихо-ботрии, расположенные на педипальпах, ногах и поверхности туловища, регистрируют коле≠бани€ воздуха. “ак называемые лировидные органы, представл€ющие собой небольшие ще≠ли в кутикуле, к перепончатому дну которых подход€т чувствительные отростки нервных клеток, €вл€ютс€ органами химического чув≠ства и служат дл€ обон€ни€. ќрганы зрени€

–ис. 398. ÷ентральна€ нервна€ систе≠ма сенокосцев (отр. Opiliones) (по
√ольмгрену): 1 Ч нервы. 2 Ч пищевод, пронизывающий
нервную массу
–ис. 399. “рахейна€
система сольпуги (из
Ћанга)
представлены простыми глазами, которые имеютс€ у большинства пау≠кообразных. ќни расположены на дорзальной поверхности головогруди и обычно их бывает несколько: 12, 8, 6, реже 2. ” скорпионов, например, имеетс€ пара срединных более крупных глаз и 2Ч5 пар боковых. ” пау≠ков чаще всего 8 глаз, расположенных обычно в две дуги, причем сред≠ние глаза передней дуги крупнее остальных.
—корпионы узнают себе подобных лишь на рассто€нии в 2Ч3 см, а некоторые пауки Ч за 20Ч30 см. ” пауков-скакунов (сем. Salticidae) зрение играет особенно важную роль: если самцам замазать глаза не≠прозрачным асфальтовым лаком, то они перестают различать самок и производить характерный дл€ периода спаривани€ Ђлюбовный танецї.
ќрганы дыхани€ Arachnida разнообразны. ” одних Ч это легочные мешки, у других Ч трахеи, у третьихЧи те и другие одновременно.
|
|
|
“олько легочные мешки имеютс€ у скорпионов, жгутоногих и у при≠митивных пауков. ” скорпионов на брюшной поверхности 3Ч6-го сег≠ментов переднебрюши€ располагаютс€ 4 пары узких щелей Ч дыхалец, которые ведут в легочные мешки (см. рис. 389). ¬ полость мешка вда-
ютс€ многочисленные параллельные друг другу листовидные складки, между которыми остаютс€ узкие щелевидные пространства, в последние проникает через дыхательную щель воздух, а в легочных листочках цир≠кулирует гемолимфа. ” жгутоногих и низших пауков имеютс€ лишь две пары легочных мешков.
” большинства других паукообразных (сольпуги, сенокосцы, лже≠скорпионы, часть клещей) органы дыхани€ представлены трахе€ми (рис. 399, 400). Ќа 1Ч2-м сегментах брюшка (у сольпуг на 1-м членике груди) имеютс€ парные дыхательные отверсти€, или стигмы. ќт каждой стигмы внутрь тела отходит пучок длинных, тонких, слепо замкнутых на концах воздухоносных трубочек эктодермального происхождени€ (обра≠зуютс€ как глубокие вп€чивани€ наружного эпители€). ” лжескорпионов и клещей эти трубочки, или трахеи, устроены просто и не ветв€тс€, у сенокосцев они образуют побочные веточки.
Ќаконец, в отр€де пауков оба рода дыхательных органов встречают≠с€ совместно. ” низших пауков, как уже отмечалось, имеютс€ только легкие; в числе 2 пар они расположены на нижней стороне брюшка. ” остальных пауков сохран€етс€ лишь одна передн€€ пара легких, а по≠зади последних имеетс€ пара трахейных пучков (рис. 400, ¬), откры≠вающихс€ наружу двум€ стигмами. Ќаконец, у одного семейства пауков (Caponiidae) легких нет вовсе, а единственные органы дыхани€ Ч 2 па≠ры трахей (рис. 400,5).
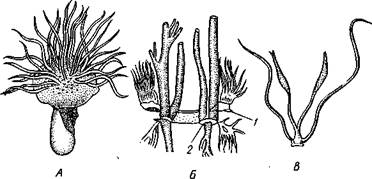
Ћегкие и трахеи паукообразных возникли независимо друг от друга. Ћегочные мешки, несомненно, более древние органы. —читаетс€, что раз-

|
| –ис. 401. —хема происхождени€ легочных мешков у паукообразных. ј Ч стади€ Ђмечехвостаї; Ѕ Ч стади€ Ђскорпионаї: / Ч дыхательна€ щель легкого. —трелки показывают токи крови и воздуха |
–ис. 400. “рахеи паукообразных (из ƒогел€). ј Ч трахеи второй па≠ры Chelifer cancroides (отр. Pseudoscopiones); Ѕ Ч трахеи паука Nops coccineus Ч две пары трахейных пучков с соответствующими стигмами (1 и 2); ¬ Ч трахеи паука Araneus diadematus
витие легких в процессе эволюции было св€зано с видоизменением брюшных жаберных конечностей, которыми обладали водные предки паукообразных и которые были похожи на жаброносные брюшные нож≠ки мечехвостов. ажда€ така€ конечность вп€тилась внутрь тела. ѕри этом образовалась полость дл€ легочных листочков (рис. 401). Ѕоковые кра€ ножки приросли к телу почти на всем прот€жении, кроме того

|
| √ |
участка, где сохранилась дыхатель≠на€ щель. Ѕрюшна€ стенка легочно-

–ис. 402. —троение сердца у пау≠кообразных (из Ћанга). ј Ч скор≠пион; Ѕ Ч паук; ¬ Ч клещ; √ Ч

