




Безусловные и условные рефлексы можно рассматривать как два «полюса», крайние типы поведенческих реакций: первые — врожденные и очень стабильные; вторые представляют собой эволюционно наиболее гибкий вариант индивидуального обучения, позволяющий сохранять информацию длительное время. Соответственно, говоря о долговременной памяти, мы подразумеваем прежде всего присущую всякой развитой ЦНС систему ее собственных условных рефлексов и ассоциаций разного уровня сложности. В целом память нервной системы можно определить как совокупность происходящих в ней процессов фиксации, сохранения и последующего воспроизведения индивидуального опыта организма.
Способность мозга к ассоциативному обучению дополняется процессами неассоциативного обучения. Они являются эволюционно более древними способами приспособления (на поведенческом уровне) к изменяющимся условиям внешней среды и уступают истинным условным рефлексам в способности к модификациям или в длительности хранения следов воздействий. Далее мы последовательно рассмотрим наиболее значимые типы неассоциативного обучения — суммацию, привыкание, долговременную потенциацию и импринтинг. Способы сохранения информации, связанные с первыми тремя, — основа нашей кратковременной памяти. Импринтинг логичнее рассматривать как вариант долговременной памяти.
| Рис. 4.13. Рефлекс втягивания жабры возникает у аплизии в ответ на прикосновение к ней или к мантийной складке: 1 — жабра; 2 — мантийная складка | 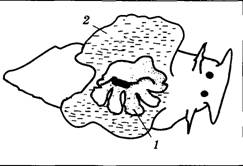
|
Отметим также, что у существ со сложно организованной нервной системой ассоциативное и неассоциативное обучение сосуществуют, причем первое в значительной мере маскирует проявления второго. Чтобы наблюдать неассоциативное обучение «в чистом виде», полезно использовать в качестве экспериментальных объектов организмы с относительно низким уровнем организации нервной системы. Именно поэтому классические работы по изучению суммации и привыкания выполнены на аплизии — крупном морском моллюске (рис. 4.13).
Суммацию можно определить как постепенное увеличение (появление) реакции при повторном предъявлении исходно индифферентного раздражителя. Суммация — простейшая форма приспособления поведения к внешним сигналам. Например, если слегка прикасаться к щупальцам гидры с интервалом в 20—30 с, то через 8—10 таких прикосновений, на которые животное исходно не реагирует, возникает реакция втягивания щупалец. Реакция эта сохраняется и при дальнейшей стимуляции. Следовательно, в нервной системе гидры произошло накопление возбуждения от отдельных раздражителей, и этот просуммированный сигнал смог запустить двигательный ответ. В нашей жизни суммация также проявляет себя достаточно часто, например, когда мы вдруг замечаем уже длительное время действовавший слабый раздражитель (капание воды из крана, легкие тактильные или болевые ощущения). Биологический смысл суммации состоит в том, чтобы «обратить внимание» нервной системы на слабые, но стабильно возникающие (или постоянно действующие) стимулы.
В основе суммации может лежать постепенное повышение чувствительности к повторяющемуся раздражителю. В этом случае адаптивные изменения происходят на входе в рефлекторные дуги различных защитных, исследовательских, пищевых и других реакций. Такой тип суммации называют сенсибилизацией. Кроме того, возможна суммация за счет снижения порога запуска реакции — фасилитация. В этом случае изменения происходят на выходе рефлекторных дуг и сенсорная специфичность раздражителя не играет заметной роли. Имея в виду описанный опыт с гидрой, можно сказать, что если суммация уже произошла, то вместо прикосновения можно включить яркий свет или оказать химическое воздействие — животное все равно будет втягивать щупальца.
Важно также то, что выработанная суммация полностью исчезает через 20—30 мин, т. е. при этом типе неассоциативного обучения длительность сохранения следов стимуляции очень невелика. Кроме того, если производить повторную суммацию, она происходит без всякого видимого облегчения и ускорения.
Механизм суммации на клеточном уровне впервые был исследован на аплизии. Измеряемой реакцией стал врожденный рефлекс втягивания моллюском жабры в ответ на сильное прикосновение. Интенсивность слабых прикосновений подбиралась так, что они исходно были незначимы, но вызывали рефлекс в результате развития суммации, т. е. после повторных применений с небольшим (до 30 с) интервалом.
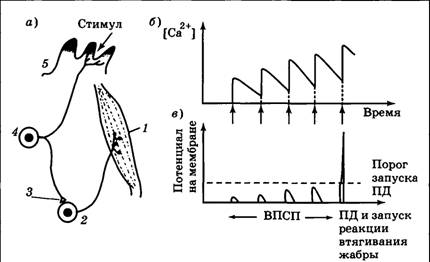
Рефлекс втягивания жабры по своей природе является пассивно-оборонительным и реализуется посредством очень короткой нервной дуги (рис. 4.14), которая состоит всего из двух клеток — сенсорной (чувствительные отростки ветвятся в ткани жабры) и двигательной (мотонейрон, управляющий мышцей, втягивающей жабру). Если внешний стимул достаточно силен, то в сенсорном нейроне возникает несколько (5— 10) потенциалов действия. В результате в синапсе между чувствительной клеткой и мотонейроном выделяется много медиатора, на мембране мотонейрона развивается большой ВПСП, а затем — ПД, который достигает мышцы, вызывая ее сокращение. При слабом стимуле возможна ситуация, когда раздражитель вызовет в сенсорном нейроне всего один ПД. Тогда медиатора в рассматриваемом синапсе выделится мало, ПД в мотонейроне не возникнет и реакция на стимул будет отсутствовать. В этом случае мы скажем, что используется исходно незначимый стимул.
Что изменится при ритмическом применении слабого стимула? Ключевым фактором, способным в итоге привести к развитию в описанной рефлекторной дуге суммации, является работа ЭПС и митохондрий пресинаптического, окончания. Напомним, что именно их нормальное функционирование обеспечивает быстрое всасывание кальция и прекращение выброса медиатора (Ca2+ попадает в пресинаптическое окончание из межклеточной среды при открывании особых каналов, реагирующих на приход ПД).
Оказывается, что всасывание кальция в ЭПС и митохондрии — длительный процесс, который может занимать десятки секунд и даже минуты. Соответственно, если внешний стимул действует достаточно часто, то белки-насосы не будут успевать удалять все ионы Ca2+ из цитоплазмы пресинаптического окончания, и кальций будет накапливаться. Чем меньше интервал между стимулами, тем быстрее идет накопление кальция. В результате при появлении очередного ПД может оказаться, что Ca2+ скопилось достаточно много для того, чтобы после выброса медиатора и генерации ВПСП потенциал действия на мотонейроне был запущен. Именно в этот момент мы увидим, что исходно незначимый стимул вызвал реакцию.
| |
| Рис. 4.14. Рефлекторная дуга, отвечающая за втягивание жабры у аплизии (а) и графики накопления ионов кальция (б) и запуска ПД в мотонейроне (в). Пять последовательных ритмических слабых воздействий на жабру отмечены стрелками (↑): 1 — мышца, оттягивающая жабру; 2— мотонейрон; 3 — синапс, в котором происходит суммация; 4 — сенсорный нейрон; 5 — жабра | |
Если не прерывать периодическую стимуляцию, то все дальнейшие раздражения будут продолжать запускать рефлекс, поскольку высокое содержание Ca2+ в пресинаптическом окончании сохраняется. Если сделать паузу, то вновь понадобится предварительная суммация (ЭПС и митохондрии удалят из цитоплазмы накопившийся кальций).
Таким образом, основной механизм развития суммации может быть определен как пресинаптическое накопление ионов Ca2+.
Тот же принцип лежит в основе ситуации, когда после однократного сильного стимула (в пресинапс входит много Ca2+) рефлекс в течение нескольких минут может запускаться исходно незначимым сигналом. Биологически это вполне оправдано: получив, например, однократное болевое подкрепление, животное еще некоторое время «соблюдает осторожность», т. е. демонстрирует пассивно-оборонительную реакцию на любые сенсорные сигналы.
Способность к суммации — распространенное свойство нервных клеток и синапсов, именно она лежит в основе более «короткого» варианта нашей кратковременной памяти. Рассматривая тот или иной зрительный объект, повторяя про себя какую-либо фразу, мы, по сути, производим ритмическую стимуляцию синапсов, включенных в передачу соответствующих сенсорных сигналов; и далее, сохраняя повышенную возбудимость в течение нескольких минут, эти синапсы «удерживают след» заинтересовавшей нас информации. Однако след этот очень быстро исчезает (по мере ослабления последствий суммации), а также легко может быть «затерт» любыми последующими сильными (значимыми) стимулами.
Суммация, видимо, явилась самым эволюционно ранним вариантом обучения. Несмотря на нестойкость происходящих изменений, она продолжает играть важную роль как один из путей индивидуальной адаптивной модификации поведения не только животных, но и человека.
Следующий тип неассоциативного обучения — привыкание. Привыкание может быть определено как ослабление (исчезновение) реакции на неоднократно предъявляемый исходно значимый раздражитель. Например, можно подобрать такую силу прикосновения к щупальцам гидры, что после 50— 70 ритмичных (с интервалом до 20—30 с) воздействий исходно проявлявший себя рефлекс втягивания щупалец исчезнет. Следовательно, в нервной системе животного каким-то образом происходит угасание реакции на повторяющийся стимул. В нашей жизни привыкание проявляется чаще всего в форме ослабления ориентировочного рефлекса, когда мы перестаем реагировать на какой-либо «надоевший» сенсорный сигнал.
Биологический смысл привыкания обусловлен стремлением нервной системы к наиболее экономному режиму функционирования.
В основе привыкания (габитуации) обычно лежат протекающие на уровне сенсорных систем процессы снижения чувствительности к повторяющемуся раздражителю. Их механизмы многообразны: адаптация чувствительных клеток, подключение к процессу обработки и проведения информации тормозных интернейронов, ослабление синаптической передачи. Первый механизм обеспечивает наиболее быстрые изменения чувствительности к постоянно действующим стимулам и рассматривается в рамках физиологии сенсорных систем.
Второй механизм является, по сути, теми же процессами суммации (пресинаптического накопления Ca2+), но протекающими на входе тормозных интернейронов. В результате повторно действующий сигнал, будучи исходно «незначимым» для тормозных нервных клеток, постепенно приводит к их включению. Как следствие, на мембранах релейных (проводящих) нейронов развиваются ТПСП и передача возбуждения к мотонейронам ослабевает (прекращается) (рис. 4.15).
Наши сенсорные системы устроены таким образом, что тормозные «фильтры», обеспечивающие привыкание, образуют несколько уровней. В результате этого происходит последовательное отсечение стабильно действующих, «неинтересных» сигналов сначала в центрах первичной обработки информации, затем в таламусе и коре.

| |
| Рис. 4.15. Схема возможных путей развития привыкания к повторяющимся сенсорным сигналам: 1 — тормозной интернейрон; 2 — мотонейрон; 3 — сенсорный нейрон | |
Ослабление синаптических процессов при привыкании также впервые показано на аплизии. Обнаружено, что избыток Ca2+ в пресинаптическом окончании, где он обязательно появится при ритмическом действии значимых стимулов, приводит к открыванию особых кальций-чувствительных К+-каналов. Выход ионов калия гиперполяризует пресинаптическое окончание и на некоторое время снижает активность электрочувствительных Ca2+-каналов, что уменьшает выброс медиатора и ослабляет синаптическую передачу.
Как и суммация, привыкание сохраняется только в течение довольно короткого времени (до 20—30 мин), а повторное привыкание идет без облегчения. В реальных условиях часто можно наблюдать, как повторяющийся слабый сенсорный сигнал сначала вызывает суммацию, а затем — привыкание. С другой стороны, можно подобрать такую низкую интенсивность стимула, что он вообще не будет вызывать реакции (даже за счет развития суммации), и такую высокую интенсивность, что привыкания к сигналу происходить не будет.
Привыкание следует отличать от утомления. В последнем случае обычно наблюдаются значительные общие изменения в состоянии рефлекторных сетей: нехватка АТФ, накопление отходов обмена веществ, истощение запасов медиаторов в пресинаптических окончаниях. Если увеличить интенсивность стимула, то эффект утомления сохраняется, а привыкание часто исчезает, поскольку усиленный сенсорный сигнал начинает использовать дополнительные нейронные цепи.
В случае аплизии показано, что привыкание, развившееся после повторных прикосновений к жабре, может быть устранено сильным воздействием на какую-либо другую часть тела. При этом активация угасшей синаптической передачи происходит за счет развития пресинаптического серотонинергического возбуждения.
Переходим к долговременной потенциации (английский эквивалент — long term potentiation — LTP).
Внешне это явление выглядит следующим образом. Вы предъявляете животному некоторый сигнал, слишком слабый для того, чтобы вызвать реакцию с первого же раза, но достаточный для возбуждения сенсорных входов. Затем после паузы в 1—2 ч (для предотвращения возможных эффектов суммации) подается сильный стимул, запускающий исследуемый рефлекс. Следующая стимуляция проводится еще через 1—2 ч с помощью слабого сигнала. При этом у существ, чья нервная система обладает способностью к LTP, возникает тот же рефлекторный ответ. Более того, интервал между сильной и последующей слабой стимуляциями может быть увеличен до 5—10 и более часов — повышенная возбудимость нервной системы продолжает регистрироваться.
По сравнению с суммацией LTP (и зафиксированная с ее помощью информация) сохраняется значительно более длительный срок, но он все же недостаточен для того, чтобы считать LTP проявлением долговременной памяти. Вот почему предлагается рассматривать LTP как вариант «длительной» кратковременной памяти, чье действие обычно распространяется на текущий период бодрствования (в случае человека — от одного ночного сна до другого).
Разберем сначала синаптическую основу долговременной потенциации, а затем попытаемся определить ее место в деятельности мозга.
В отличие от суммации, LTP происходит на строго определенных синапсах: во-первых, глутаматергических и, во-вторых, снабженных несколькими типами рецепторов к глутаминовой кислоте, основным из которых должен быть NMDA-тип. Схема такого синапса представлена на рис. 4.16. Напомним, что NMDA-рецепторы обладают уникальным свойством находиться в двух способных переходить друг в друга состояниях — рабочем и инактивированном. В последнем случае канал NMDA-рецептора оказывается перекрыт ионом Mg2+ (магниевая пробка).

| |
| Рис. 4.16. Схема синапса, способного к развитию долговременной потенциации: 1 — метаботропный (не-NMDA-peцептор); 2 — NMDA-рецептор, находящийся в нерабочем состоянии; 3 — везикула с глутаматом; 4 — хемочувствительный ионный канал | |
Рассмотрим работу синапса, способного к LTP. Его исходное состояние характеризуется наличием только инактивированных NMDA-рецепторов. Если подать на такой синапс слабый стимул, то, как и в случае суммации, возникает лишь одиночный ПД, приводящий к выбросу небольшого количества глутаминовой кислоты. При этом медиатор может вызвать открывание Na+-каналов, связанных только с не-NMDA-peцепторами. В результате ВПСП, развивающийся на постсинаптической мембране, будет мал, ПД в следующей клетке не возникнет и реакция на слабый стимул наблюдаться не будет.
Применение сильного стимула вызовет выделение большого количества глутаминовой кислоты, и возбуждения даже He-NMDA-рецепторов окажется достаточно, чтобы на постсинаптической мембране возник ПД, а на уровне всего организма — поведенческий ответ. Параллельно происходит ключевое событие, запускающее LTP: ПД, являясь скачком потенциала на постсинаптической мембране, провоцирует выход ионов Mg2+ из каналов NMDA-рецепторов в межклеточную среду — «выбивание магниевых пробок» переводит NMDA-рецепторы в рабочее состояние.
Возврат пробок осуществляется постепенно — в течение нескольких часов, и все это время синапс остается в состоянии повышенной готовности к проведению информации (в этом смысл термина «потенциация»). Отсюда следует, что, если сейчас посредством слабой стимуляции подать на вход синапса даже одиночный ПД, выделяющаяся глутаминовая кислота подействует на все рецепторы (в том числе на NMDA-тип). Поэтому, несмотря на небольшое количество медиатора, развивающийся на постсинаптической мембране ВПСП окажется достаточно велик для последующей генерации ПД и появления поведенческого ответа.
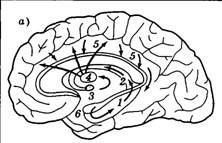
Наиболее плотная концентрация NMDA-рецепторов в мозге высших позвоночных обнаружена на входе в гиппокамп — ключевую структуру круга Папеца (рис. 4.17, а). Этот круг — важнейший структурно-функциональный комплекс переднего мозга и в качестве основных структур включает: собственно гиппокамп, выходящий из него свод, мамиллярные тела, передние ядра таламуса, ряд структур коры (важнейшая из них — поясная извилина) и зубчатую извилину, чьи аксоны заканчиваются в гиппокампе. Необычной и характерной чертой данного комплекса является его цикличность, замкнутость на свои собственные ядра. Этот факт еще в 60-е годы позволил сформулировать гипотезу о круге Папеца как месте хранения циркулирующих по замкнутому контуру нервных сигналов. В 90-е годы это представление было дополнено данными об NMDA-рецепторах, что позволило сформировать представление о LTP как одном из физиологических механизмов памяти.
|
|
| Рис. 4.17. Круг Папеца (а) и его упрощенная структура (б): 1 — гиппокамп; 2 — свод; 3 — мамиллярное тело; 4 — передние ядра таламуса; 5 — поясная извилина; 6 — зубчатая извилина; • — синапсы, содержащие NMDA-рецепторы |

Рассмотрим схему, приведенную на рис. 4.17, б. На ней из всего множества структур круга Папеца оставлены лишь две ключевые — гиппокамп, обладающий NMDA-рецепторами, и поясная извилина, для которых характерны множественные связи с самыми разными отделами коры и подкорки (в том числе с гипоталамусом и центрами бодрствования). Гиппокамп и поясная извилина соединены множественными взаимными, формирующими круг связями (волокна свода и др.). Как происходит фиксация информации с помощью этой системы?
Представим себе человека ранним утром, его мозг полностью отдохнул. Можно считать, что при этом структуры круга Папеца находятся в «очищенном» состоянии, все пути передачи информации пока закрыты и магниевые пробки располагаются на своих местах — в каналах NMDA-рецепторов. Далее начинается обычный рабочий день: человек встает, умывается, завтракает, совершая обычный утренний ритуал.
Если при этом ничего необычного не происходит, то уровень возбуждения в мозге невелик и сохранение памяти идет только за счет самых нестойких суммационных механизмов. Однако если происходит что-то необычное, значимое, имеющее эмоциональную окраску, то уровень активации в соответствующих сенсорных и ассоциативных центрах коры резко повышается. Это приводит к возбуждению поясной извилины, переадресовке сигналов на вход гиппокамп, и, как в случае применения сильного стимула, «выбиванию магниевых пробок» в соответствующих стимулу или ситуации информационных каналах.
Момент выбивания Mg+-пробок приводит к замыканию информационного контура в круге Папеца, по которому затем будет происходить циркуляция возбуждения (источником слабой активации являются центры бодрствования). И пока контур функционирует — информация о прошедшем значимом событии будет периодически возвращаться через поясную извилину в кору, «напоминая» о себе.
Из описанной схемы следует несколько выводов. Во-первых, легко понять, что емкость (объем памяти) круга Папеца хотя и велика, но не безгранична. Каждый из нас может вспомнить ощущения, возникающие тогда, когда этот блок памяти переполняется — при просмотре нескольких кинофильмов подряд, посещении крупного музея, многочасовом сидении на лекциях.
Во-вторых, система круга Папеца и LTP оказываются чувствительными к электрошоковым воздействиям, когда тотальное выбивание магниевых пробок стирает с нейронных контуров текущую информацию. В результате человек может забыть о событиях, имевших место за несколько часов до электрошока (ретроградная амнезия).
В-третьих, восстановление ресурса этой системы, ее очистка и возврат магниевых пробок на место требует достаточно длительного прекращения циркуляции возбуждения по соответствующим контурам, что возможно лишь при выключении центров бодрствования и засыпании. При этом информация, записанная на контурах круга Папеца, частично теряется, а частично переходит в долговременную форму хранения, о чем будет сказано ниже.
Отметим также, что синапсы с NMDA-рецепторами встречаются не только в гиппокампе, но и в других зонах ЦНС — сенсорных центрах, двигательных центрах, ассоциативной коре больших полушарий, где они выполняют аналогичную функцию — обеспечивают быстрое формирование каналов для передачи приоритетной информации, переключают потоки сигналов с канала на канал (своеобразные «стрелочники»).
Последний из рассматриваемых видов неассоциативного обучения — импринтинг, или запечатление. Импринтинг определен как формирование в некоторые периоды онтогенеза устойчивой индивидуальной избирательности по отношению к внешним стимулам.
Можно выделить три наиболее показательных варианта импринтинга: запечатление родителя детенышем; запечатление детеныша родителем и импринтинг будущего полового партнера. Первый из них подробно описан и изучен К. Лоренцом. В его работах показано, что птенцы выводковых птиц, реализуя врожденное детское поведение, следуют за первым движущимся объектом, который увидели после рождения; то же относится и к ряду млекопитающих. Если такой объект окажется не матерью, а человеком, неживым предметом (что и проделывалось в ходе экспериментов), птенцы также будут двигаться за ним. Критический период для запечатления родителя — первые часы (максимум — дни) жизни детеныша.
Во втором случае можно наблюдать возникновение у матери импринтинга на детеныша. Так, коза в течение примерно часа после родов имеет особую чувствительность к запаху козленка. В дальнейшем именно он будет восприниматься ею как собственный детеныш, а остальные козлята будут отвергаться. У самок колониальных птиц и тюленей может происходить запечатление голоса детеныша.
Третий вариант демонстрирует, что, кроме взаимоотношений родитель — детеныш, импринтинг имеет особое значение для формирования полового поведения и предпочтения будущего полового партнера. Это доказано путем замены родителей у голубей, кур, уток. В экспериментах характерно окрашенные родители одной породы воспитывали вместе с собственными детенышами птенца другой породы. Когда такой птенец вырастал, то предпочитал спариваться с птицами, имеющими такую же окраску, как и его приемные родители. Критический период для запечатления будущего полового партнера — стадия окончательного полового созревания птенцов, когда происходит запечатление внешнего вида как родителей, так и соседей по гнезду (братьев и сестер). Запечатление это сказывается и на других компонентах зоосоциального поведения.
В целом можно сказать, что импринтинг связан с установлением жизненно важных внутрисемейных, внутригрупповых и внутривидовых контактов, при этом по своей сути поведенческие реакции, связанные с импринтингом, являются врожденными. Однако, в отличие от обычных безусловных рефлексов, в данном случае стимул, запускающий реакцию, исходно не определен. В ходе запечатления происходит запоминание характеристик этого стимула (внешний вид, запах родителя, детеныша, будущего полового партнера) и устанавливается связь между сенсорными центрами и центром соответствующей врожденно запрограммированной реакции. В отличие от условного рефлекса, эта связь, во-первых, образуется только в строго определенный период жизни животного; во-вторых, образуется без подкрепления; в-третьих, в дальнейшем оказывается очень стабильной, практически не подлежит угашению и может сохраняться в течение всей жизни особи.
На рис. 4.18 показано, что обучающийся нейрон, способный запустить некоторую врожденную реакцию организма, исходно получает большое количество пока еще незначимых сенсорных сигналов, причем каждый — по своему каналу. В критический период онтогенеза происходит выбор самого активного сенсорного входа и соответствующий синапс резко увеличивает свою эффективность. В результате в обучающемся нейроне возникают ПД, включающие определенную поведенческую программу.

| |
| Рис. 4.18. Принципиальная схема механизма, связанного с формированием импринтинга: 1 — обучающийся синапс; 2 — обучающийся нейрон; 3 — двигательный нейрон | |
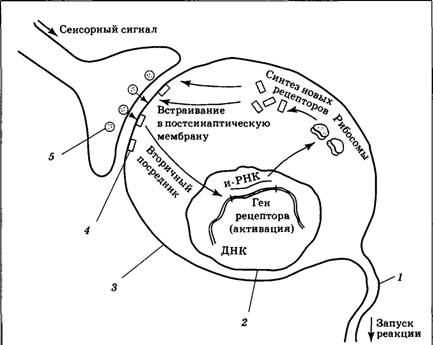
Понятно, что ключевую роль в данном процессе играет механизм модификации обучающегося синапса. Каким образом он начинает работать более эффективно, т. е. генерировать больший ВПСП в ответ на то же количество медиатора? Современные представления о механизмах этого процесса представлены на рис. 4.19, где показано, что выделяющийся из пресинаптического окончания медиатор (глутаминовая кислота) действует на соответствующие метаботропные рецепторы, запуская образование вторичного посредника (например, ц‑АМФ). Вторичный посредник, передавая сигнал внутри обучающегося нейрона, проникает в ядро и воздействует на ДНК. Конечным результатом такого воздействия является активация генов, управляющих синтезом рецепторов к глутаминовой кислоте: с них «снимается копия» — и-РНК, которая поступает к рибосомам. Рибосомы синтезируют новые белки-рецепторы к глутаминовой кислоте, которые встраиваются в постсинаптическую мембрану самого активного синапса, передающего информацию о характеристиках импринтингуемого объекта.
|
| Рис. 4.19. Схема механизма импринтинга на молекулярном уровне: 1 — аксон; 2 — ядерная оболочка; 3 — мембрана обучающегося нейрона; 4 — рецептор к медиатору; 5 — везикулы с медиатором |
Встраивание в постсинаптическую мембрану дополнительных рецепторов является тем ключевым фактором, который приводит к увеличению эффективности синаптической передачи. Как только рецепторов окажется достаточно много, ВПСП на мембране обучающегося нейрона достигнет критического уровня, и ПД (поведенческая реакция) будет запущена. Внешне это выглядит так: детеныш будет рассматривать находящиеся перед ним объекты, пока наконец один из них, привлекший особое внимание за счет движений или звуков, не начнет запускать различные компоненты детского поведения.
Специально подчеркнем, что описанные нейрохимические и синаптические изменения протекают не мгновенно, а требуют времени, т. е. для успешного импринтинга важен фактор стабильного сенсорного «давления» на обучающийся нейрон (например, постоянное присутствие матери). Если это условие не будет выполнено, критический для запечатления период может пройти и импринтинг вообще не возникнет.
Важно также то, что в дальнейшем обучающийся нейрон продолжает поддерживать концентрацию рецепторов на постсинаптической мембране «запечатленного» синапса на том же высоком уровне. Именно это свойство обучающихся нейронов обеспечивает чрезвычайную стабильность импринтинга, позволяющую рассматривать его как специфический вариант долговременной памяти.
В чем биологический смысл импринтинга и почему эволюции понадобилось создавать что-то промежуточное между условными и безусловными рефлексами? Дело в том, что обучение, происходящее по типу импринтинга, обслуживает самые ключевые моменты зоосоциального поведения. Эти моменты настолько важны (отношения детеныш — родитель, половое поведение), что формировать их на базе условных рефлексов, способных при определенных условиях угасать, блокироваться, видимо, нецелесообразно. С другой стороны, реализовать такое поведение на основе безусловных рефлексов тоже не всегда возможно, поскольку объект, на который направлено поведение, может иметь нестабильные (разный запах) или очень «тонкие» (особенности расцветки оперения) сенсорные свойства. Таким образом, импринтинг оказывается очень полезен в тех специфических ситуациях, когда «чувствительности» безусловных рефлексов не хватает, а относительная нестабильность условных рефлексов может поставить жизнь или воспроизводство особи под угрозу.
Приведем поясняющий пример. Как известно, у самцов уток яркая и видоспецифичная окраска; у противоположного пола окраска более монотонная, и отличить самок разных видов непросто. Оказывается, что различение самками «своих» самцов обусловлено врожденной поведенческой программой, а вот различение самцами самок требует импринтинга. Доказано это в уже упоминавшихся экспериментах с подменой птенцов: если самку выращивать в гнезде другого вида, то она, несмотря ни на что, в качестве полового партнера будет выбирать самца своего вида. Если же в чужом гнезде выращен самец, то его половое поведение будет адресоваться самкам, похожим на приемную мать.
Не исключено, что импринтинг присутствует (в качестве компонента, хотя бы частично определяющего обучение) в большинстве ситуаций, когда та или иная безусловно-рефлекторная система срабатывает впервые. В таком случае становится более понятным значение ранних детских впечатлений, которые могут на долгие годы (и даже на всю жизнь) запрограммировать наши вкусовые и запаховые предпочтения, особенности оборонительного поведения и т. п. Поиск таких впечатлений, их анализ, «поднятие» из сферы бессознательного лежит в основе многих психотерапевтических методик.
Итак, импринтинг является специфическим вариантом длительного сохранения мозгом информации. Однако основной тип долговременной памяти связан все же с ассоциативным обучением.
На рис. 4.20 приведена структура нейрональной системы, обеспечивающей формирование временной связи. Этот рисунок отличается от аналогичного, относящегося к импринтингу, прежде всего присутствием фактора положительного подкрепления. В случае условного рефлекса на обучающемся нейроне сходятся два потока сигналов — сенсорная информация от центра, обеспечивающего восприятие исходно незначимого стимула, и сигналы от гипоталамической системы положительного подкрепления. На рис. 4.21 показано, как такая конвергенция выглядит на синаптическом уровне. При этом сенсорные проекции в качестве медиатора используют все ту же глутаминовую кислоту (рецепторы помечены прямоугольниками), системы подкрепления — чаще всего моноамины (серотонин, норадреналин, дофамин; рецепторы помечены треугольниками).

| |
| Рис. 4.20. Принципиальная схема механизмов, связанных с ассоциативным обучением (формированием условных рефлексов): 1 — обучающийся синапс; 2 — обучающийся нейрон; 3 — двигательный нейрон; 4 — сенсорный нейрон | |

| |
| Рис. 4.21. Схема механизма ассоциативного обучения на молекулярном уровне: 1 — аксон; 2 — моноамины и рецепторы к ним; 3 — глутаминовая кислота и рецепторы к ней | |
В отличие от импринтинга, где для осуществления обучения было достаточно постоянно действующего сенсорного притока, в случае условного рефлекса необходима одномоментная активация обоих синаптических входов. В этом случае в цитоплазме одновременно появятся два вторичных посредника (на схеме ВтП1 ВтП2), которые совместно подействуют на ядерную ДНК. Дальнейшие последствия аналогичны описанным для импринтинга: активируется синтез рецепторов к глутаминовой кислоте, которые затем встраиваются в постсинаптическую мембрану самого активного сенсорного синапса (подготовка мембраны к встраиванию дополнительных рецепторов является, видно, одной из функций ВтШ). В результате этого эффективность синаптической передачи резко возрастает, и ранее незначимый сигнал начинает запускать реакцию. Внешне это выглядит, как, например, появление реакции слюноотделения в ответ на включение лампочки.
Давно известно, что формирование долговременной памяти сопровождается синтезом РНК и белков. В 60-е годы это привело к появлению представлений о «молекулах памяти» и надежде на то, что когда-нибудь будут найдены и расшифрованы биополимеры, кодирующие индивидуальную память. Но все оказалось гораздо сложнее. Молекул памяти как носителей личного опыта индивидуума нет, и в этом отличие нервной памяти от генетической. Долговременная память связана с множественными изменениями свойств синапсов в нейронных сетях коры, которые при этом не имеют стандартной локализации. В случае каждого условного рефлекса мы знаем начало временной связи (сенсорный центр), ее окончание (корковое представительство безусловного рефлекса), но какова ее конкретная «траектория», какие нейроны и синапсы из миллиардов возможных претендентов оказались задействованы — остается тайной.
И еще одно существенное замечание. В случае импринтинга мы видели, что для успешного запечатления необходимо длительное сенсорное «давление» на обучающийся нейрон. Но если механизмы синаптических изменений при импринтинге и ассоциативном обучении близки, то каким же образом обычно очень непродолжительно действующий условный стимул ухитряется «раскачать» ядерную ДНК и рибосомальную активность? Ответ может быть получен за счет привлечения к процессу формирования долговременной памяти систем кратковременной памяти и круга Папеца.
Если сенсорный стимул сопровождается подкреплением, то это «подтверждает» его значимость и вызывает изменения в круге Папеца (активация NMDА-рецепторов в соответствующих нейронных циклах). Затем, когда действие условного стимула прекратится, активация из круга Папеца может поступать к обучающимся нейронам коры, «подменяя» реальный сигнал и продолжая активировать нейрохимические механизмы долговременного запоминания.
Изложенная концепция позволяет объяснить последствия, например, двустороннего повреждения круга Папеца, когда страдает не только система кратковременной памяти, но и формирование новой долговременной памяти. При этом уже имевшаяся у пациента долговременная память не нарушается. Таким образом, еще одну функцию круга Папеца и основной его структуры гиппокампа можно определить как обеспечение перезаписи информации из кратковременной формы хранения в долговременную.
Завершая главу, отметим, что в целом описанные типы высшей нервной деятельности можно свести к четырем основным вариантам:
1) безусловные рефлексы — в этом случае нервные связи заданы врожденно, они очень устойчивы;
2) суммация, привыкание, долговременная потенциация — нервные связи, сохраняющие информацию, являются врожденными (круг Папеца), но функционально они неустойчивы и могут в течение короткого времени как усиливаться, так и ослабляться;
3) импринтинг — нервная связь формируется в результате обучения, в дальнейшем она очень устойчива и не исчезает;
4) ассоциативное обучение — нервная связь формируется в результате сочетания с подкреплением; в дальнейшем при определенных условиях она может укрепляться либо угасать (условное торможение).

