




Содержание
Введение -3
Основные виды модификаций-3
Ограниченный протеолиз-4
Ковалентные модификации-5
Включение кофакторов-7
Ассоциация в олигомеры-8
Список источников-9
Введение
Полипептидные цепи могут подвергаться структурным модификациям, либо будучи ещё связанными с рибосомами, либо после завершения синтеза. Эти конформационные и структурные изменения полипептидных цепей получили название посттрансляционных изменений.
Многие модификации осуществляются в ЭР. Здесь происходят фоддинг полипептидных цепей и формирование уникальной третичной или четвертичной структуры белков.
Основные виды модификаций:
1. Ограниченный протеолиз
2. Ковалентная модификация
3. Включение Ко-факторов
4. Ассоциация в олигомеры

Ограниченный протеолиз
Многие белки, секретируемые из клеток, первоначально синтезируются в виде молекул-предшественников, функционально неактивных. Удаление части полипептидной цепи специфическими эндопротеазами (гидролазы –расщепляют пептидные связи в молекуле белка) приводит к образованию активных молекул. Некоторые белки-предшественники расщепляются в ЭР или аппарате Гольджи, другие - после секреции. Так, неактивные предшественники секретируемых ферментов – зимогены (проферменты) - образуют активный фермент после расщепления по определённым участкам молекулы: зимоген панкреатической железы трипсиноген превращается в активный трипсин после секреции в тонкий кишечник.

Иногда процесс созревания идет вместе с процессом транспорта. Наглядным примером последовательного двухстадийного протеолиза служит образование активных форм пептидных гормонов (например, инсулина или глюкагона) из препрогормонов. Первоначально, N-концевой сигнальный пептид (Сигнальный пептид, или сигнальная последовательность, — короткая (от 3 до 60 аминокислот) аминокислотная последовательность в составе белка, которая обеспечивает посттрансляционный транспорт белка в соответствующую органеллу (ядро, митохондрия, эндоплазматический ретикулум, хлоропласт, апопласт или пероксисома). После доставки белка в органеллу сигнальный пептид может отщепляться под действием специфической сигнальной протеазы.) молекулы-предшественника удаляется в ЭР в процессе синтеза белка и образуется неактивный прогормон. Затем прогормон в секреторных гранулах, формирующихся в аппарате Гольджи, подвергается действию эндо- и/или экзопротеаз и превращается в активный гормон.
Типы сигнальных последовательностей
| Органелла | Аминокислотная последовательность |
| клеточное ядро | -Pro-Pro-Lys-Lys-Lys-Arg-Lys-Val- |
| транспортировка к эндоплазматическому ретикулуму | H2N-Met-Met-Ser-Phe-Val-Ser-Leu-Leu-Leu-Val-Gly-Ile-Leu-Phe-Trp-Ala-Thr-Glu-Ala-Glu-Gln-Leu-Thr-Lys-Cys-Glu-Val-Phe-Gln- |
| задержка в эндоплазматическом ретикулуме | -Pro-Pro-Lys-Lys-Lys-Arg-Lys-Val- |
| митохондрия (матрикс) | H2N-Met-Leu-Ser-Leu-Arg-Gln-Ser-Ile-Arg-Phe-Phe-Lys-Pro-Ala-Thr-Arg-Thr-Leu-Cys-Ser-Ser-Arg-Tyr-Leu-Leu- |
| пероксисома (PTS1) | -Ser-Lys-Leu-COOH |
| пероксисома (PTS2) | H2N-----Arg-Leu-X5-His-Leu- |
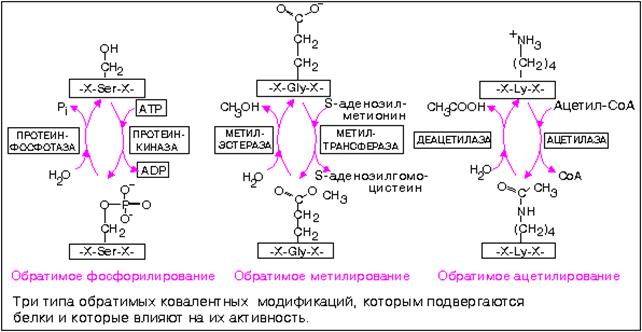
Ковалентная модификация
Структурные белки и ферменты могут активироваться или инактивироваться в результате присоединения различных химических групп: фосфатных, ацильных, метальных, олигосахаридных и некоторых других.
Фосфорилирование белков осуществляется по гидроксильным группам серина, треонина итирозина ферментами из группы протеинкиназ, тогда как дефосфорилирование катализируют гидролитические ферменты фосфопротеинфосфатазы.
Метилирование белка процесс посттрансляционной модификации белка, заключающийся в присоединении метильной группы к аминокислотам полипептида. Метилирование осуществляется главным образом по аминокислотным остаткам лизина, аргинина и гистидина (N-метилирование), а также по остаткам глутаминовой и аспарагиновой кислот (О-метилирование).
Гликозилированием называют процесс присоединения к полипептидной цепи различных углеводных остатков. Все секретируемые белки эукариот бывают в той или иной степени гликозилированы.
N-гликозилирование осуществляется по аспарагину, расположенному через один аминокислотный остаток от триптофана, и происходит постадийно.

В процессе О-гликозилирования происходит присоединение одного-двух углеводных остатков преимущественно по серину и триптофану. Белки, входящие в состав плазматических мембран или секретирующиеся из клеток, подвергаются гликозилированию. Углеводные цепи присоединяются то гидроксильным группам серина или треонина (О-гликозилирование). Последовательное наращивание углеводного фрагмента происходит в ЭР и аппарате Гольджи.

Включение Кофакторов
Большинство ферментов для проявления ферментативной активности нуждается в низкомолекулярных органических соединениях небелковой природы и/или в ионах металлов. Более 25% всех ферментов для проявления полной каталитической активности нуждается в ионах металлов. Рассмотрим роль кофакторов в ферментативном катализе.
Кофактор + белковая часть фермента – холофермент.
Кофакторы неорганической природы – ионы металлов.
Функции:
1. Поддерживают конформацию белковой молекулы
2. Мостик межу белковой и небелковой частью
3. Может связывать субстрат и фермент
4. Может участвовать в катализе
Кофакторы органической природы – витамины, нуклеотиды, витаминоподобные в-ва.
Функции: переносят функциональные группы.
Роль металлов в присоединении субстрата
в активном центре фермента
Ионы металла выполняют функцию стабилизаторов молекулы субстрата, активного центра фермента и конформации белковой молекулы фермента, а именно третичной и четвертичной структур.
Ионы металлов - стабилизаторы молекулы субстрата
Для некоторых ферментов субстратом служит комплекс превращаемого вещества с ионом металла. Например, для большинства киназ в качестве одного из субстратов выступает не молекула АТФ, а комплекс Mg2+-ATФ. В этом случае ион Mg2+ не взаимодействует непосредственно с ферментом, а участвует в стабилизации молекулы АТФ и нейтрализации отрицательного заряда субстрата, что облегчает его присоединение к активному центру фермента.

Ассоциация в олигомеры
1. Многие белки имеют в своем составе несколько полипептидных цепей. Такие белки называют олигомерными, а отдельные цепи — протомерами.
Протомеры в олигомерном белке соединены множеством слабых, нековалентных связей (гидрофобных, ионных, водородных).
Взаимодействие протомеров осуществляется благодаря комплементарности их контактирующих поверхностей.
Количество протомеров в белках может сильно варьировать: гемоглобин содержит 4 протомера.
2. Взаимодействие одного протомера с другими можно рассматривать как частный случай взаимодействия белка с лигандом. Каждый протомер служит лигандом для других протомеров.
3. Количество и порядок соединения протомеров в белке называется четвертичной структурой.

