




Схема дихал ланц мітохон

47.. Хеміосмотична теорія Мітчелла (теорія окисного фосфорилювання)
Пояснення механізмів спряження роботи дихального ланцюга та синтезу АТФ було запропоноване англійським біохіміком Пітером Мітчеллом у 1961 році, яке потім знайшло експериментальне підтвердження у роботах багатьох дослідни-ків. За винайдення механізмів окисного фосфорилювання П. Мітчелл у 1978 році отримав Нобелівську премії в галузі хімії («За внесок у розуміння процесу перенесення біологічної енергії, який зроблений завдяки створенню хеміосмотичної теорії»).

Рисунок 19 - Спряження роботи дихального ланцюга із синтезом АТФ (за теорією П.Мітчелла)
Основними постулатами теорії Мітчелла є таке (рис. 19):
1) внутрішня мембрана мітохондрій непроникна для іонів та малих молекул (за винятком молекул води);
2) дихальний ланцюг працює як «помпа», що викачує протони з матриксу в міжмембранний простір – рух 2 електронів від субстрату на кисень призводить до перенесення 8-10 Н+ (протони транспортуються через I, III та IV комплекси) через мембрану;
3) робота дихального ланцюга створює електрохімічний градієнт протонів (∆μН+), оскільки вони вільно через внутрішню мітохондріальну мембрану повернутися в матрикс не можуть і будуть накопичуватись у міжмемб-ранному просторі; ∆μН+ - це проміжна форма зберігання енергії окиснення субстратів;
4) енергію протонного градієнта використовує Н+-АТФ- синтаза (V комплекс) для синтезу АТФ, коли через одну з її субодиниць протони повертаються в матрикс;
5) існують сполуки - роз’єднувачі окисного фосфорилю-вання, які порушують електрохімічний градієнт протонів і знижують ефективність роботи Н+-АТФ-синтази.
На рис. 20 наведена структура протонної АТФ-синтази (Н+-АТФ-синтази, Н+-АТФ-аза), яка безпосередньо використовує енергію протонного градієнта, що був створений дихальним ланцюгом, для синтезу АТФ.
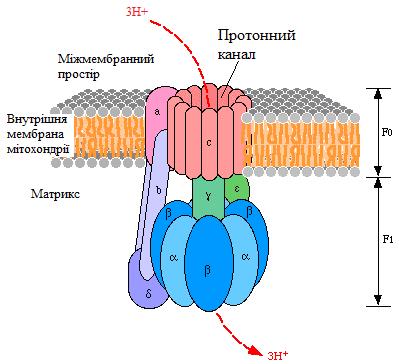
Рисунок 20 - Структура Н+-АТФ-синтази
Протонна АТФ-синтаза – це олігомерний білок, який вмонтований у внутрішню мембрану мітохондрії і за будовою нагадує гриб (рис. 19). Вона містить дві субодиниці:
Fo – протонний канал (о - від «олігоміцин»); лише через цей канал протони можуть повернутися в матрикс;
F1 – фермент, який використовує енергію, що вивільняється при транспорті протонів через Fo для синтезу АТФ з АДФ та Фн.
П.Мітчелл у своїй теорії теоретично віддав функцію спряження окиснення та фосфорилювання саме Н+-АТФ-азі. Експериментальне піддтердження цей факт знайшов у працях Джона Уокера та Пола Бойера, які за «З’ясування ензимного механізму, що лежить в основі синтезу аденозин-фосфату» в 1997 році отримали Нобелівську премію з хімії.
На сьогодні відомо, що при транспорті протонів через Fo-субодиницю відбуваються конфірмаційні зміни в активному центрі F1-субодиниці, які призводять до її активації і відповідно до синтезу АТФ та її вивільнення. Молекули АТФ, що синтезуються, транспортуються у цитозоль за допомогою транслокази.
Для синтезу молекули АТФ, її вивільнення та транспорту у цитозоль потрібна енергія 4 протонів (40% цієї енергії іде на синтез АТФ, 60% виділяється у вигляді тепла).
48. Кількість молекул неорганічного фосфату, яке перейшло у зв’язану форму (тобто у АТФ) у перерахунку на один атом кисню має назву коефіцієнт окисного фосфорилювання і позначається Р/O (коефіцієнт фосфорилювання).
Коефіцієнт Р/O чисельно дорівнює кількості молекул АТФ, що синтезувалися в результаті транспорту 2 ē на один атом кисню. Тому для субстратів, що окислюються під дією НАД-залежних дегідрогеназ Р/O= 3 (наприклад, для пірувату, α-кетоглутарату, ізоцитрату, малату). Для субстратів, які окислюються за допомогою ФАД-залежних дегідрогеназ, цей коефіцієнт дорівнює 2 (наприклад, для сукцинату, ацил-КоА, гліцерил-3-фосфату).
Цікаво
Бура жирова тканина добре розвинена у новонарод-жених дітей. У дорослих вона залишається в незначній кількості на шиї, між лопатками, навколо нирок, щитоподібної залози та аорти. Бура жирова тканина містить велику кількість мітохондрій, цитохроми яких надають тканині коричневого кольору. Мітохондрії цієї жирової тканини мають окислювальну здатність, що у 20 разів більша, ніж мітохондрій білої жирової тканини.
Для дітей ця тканина має велике значення, оскільки невеликий за розмірами організм дитини здатний дуже швидко втрачати тепло. Кількість активного бурого жиру у дорослих різна і залежить від ваги. Як правило, у гладких людей кількість бурої жирової тканини менша. Організм дорослих, які мають буру жирову тканину, відрізняється підвищеною швидкістю метаболізму, оскільки, як відомо, 50 грамів бурого жиру за добу допомагає додатково «спалювати» 500 ккал енергії.
Інгібітори дихального ланцюга
Робота дихального ланцюга може бути інгібована під дією різноманітних клітинних отрут, які взаємодіють з білками-переносниками та порушують транспорт електронів:
- Інгібітори I комплексу — барбітурати (амітил, секонал), ротенон, пієрицидин А.
- Інгібітор II комплексу — малонат.
- Інгібітор III комплексу — антиміцин А.
- Інгібітори IV комплексу — Н2S, цианіди, СО.
Наявність у клітині вказаних інгібіторів призводить до розвитку гіпоенергетичного стану, що, в свою чергу, в деяких випадках може призвести до загибелі клітини.
Деякі речовини роз’єднують окиснення та фосфорилювання, тому мають назву роз’єднувачів окисного фосфорилювання. Ці сполуки ліпофільні, легко дифундують через внутрішню мембрану мітохондрій, можуть зв’язувати протони та переносити їх у матрикс за градієнтом концен-трацій Н+ (тому вони належать до сполук протонофорів).
У разі наявності в клітині роз’єднувача окиснення субстратів відбувається дуже активно, дихальний ланцюг мітохондрій працює і намагається створити електрохімічний градієнт протонів, але цього не відбувається. Пояснення в тому, що молекули роз’єднувача захоплюють протони з міжмемб-ранного простору і транспортують їх у матрикс. За таких умов окиснення субстратів не приводить до створення протонного градієнта і, відповідно, до синтезу АТФ. Енергія окиснення перетворюється в теплову енергію, тобто коефіцієнт окисного фосфорилювання знижується і спостерігається виділення тепла (пірогенна дія).
Класичним прикладом роз’єднувача є 2,4-динітрофенол:

Крім 2,4-динітрофенолу до синтетичних роз’єднувачів потрібно віднести деякі похідні вітаміну К, анестетики, антибіотики валіноміцин та граміцидин, ацетилсаліцилову кислоту, гербіциди. До природних роз’єднувачів належать продукти перикисного окиснення ліпідів, довголанцюгові жирні кислоти, великі дози териоїдних гормонів.
Саме тому в людей, які хворіють на гіпертиреоз спостерігається підвищення температури тіла, активне окиснення субстратів і тому, незважаючи на підвищений апетит, вони втрачають вагу.
Дихальний контроль
Процес синтезу АТФ залежить від енергетичного статусу клітини, який, у свою чергу знаходить своє вираження у кількісному співвідношенні АТФ/АДФ. При зростанні потреб клітини в енергії підвищується концентрація АДФ і значення цього співвідношеня знижується. Це супроводжується активацією тканинного дихання і, відповідно, окисного фосфорилювання.
Процес залежності дихання клітин від вмісту аденілових нуклеотидів був вивчений in vitro. Результати дослідження показали, що за наявності в середовищі всіх компонентів, за винятком АДФ, поглинання кисню клітиною не відбувається. Внесення в інкубаційну суміш АДФ стимулює дихання, активність якого знижується при зростанні концентрації АТФ. Швидкість дихання знижується до нуля, коли вся АДФ перетворюється в АТФ. Тому залежність роботи дихального ланцюга від співвідношення АТФ/АДФ отримала назву дихальний контроль. Механізм дихального контролю відрізняється високою точністю і швидко реагує навіть на невеликі зміни концентрацій АТФ та АДФ. Незначне підвищення концентрації АДФ призводить до активації дихання та фосфорилювання.
Зниження синтезу АТФ призводить до розвитку гіпоенергетичного стану в організмі. Синтез АТФ знижується при:
1) гіпоксії;
2) голодуванні, дефіциті деяких вітамінів (В1, В2, РР) та мінералів (Fe, Cu);
3) наявності інгібіторів дихального ланцюга та інгібіторів окисного фосфорилювання;
4) наявності роз’єднувачів окиснення та фосфорилювання.
АТФ не депонується в клітинах організму, постійно відбу-вається синтез та розпад молекул цього нуклеозидтрифосфату. Загальний вміст АТФ в організмі - 30-50 г, але час «життя» молекули АТФ – менше 1 хв. За добу в організмі людини синтезується та розщеплюється 40-60 кг АТФ.
50. Окиснення, яке відбувається за участі цитохрому Р450, має назву мікросомальне окиснення.
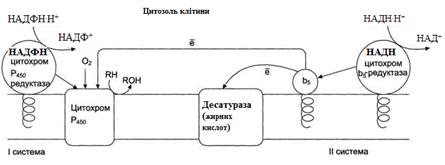
Рисунок 21 – Електрон-транспортні ланцюги мікросомального окиснення: I система – НАДФН-цитохром Р450 редуктазна;
II система – НАДН-цитохром b5 редуктазна.
Мікросомальне окиснення відбувається за участі електрон-траспортних ланцюгів, які містять білки-переносники, що вбудовані в мембрани. Існують два ланцюги (на рис. 20 – системи):
I – до складу входить два ферменти: НАДФН ⋅ Н+-цитохром Р450 редуктаза та цитохром Р450. НАДФН ⋅ Н+-цитохром Р450 редуктаза містить два коферменти ФАД та ФМН, які беруть участь у транспорті електронів з НАДФН ⋅ Н+ на цито-хром Р450 (рис. 22). Цитохром Р450 каталізує гідроксилювання субстратів.
II – містить фермент НАДН-цитохром b5 редуктазу та цитохром b5. Коферментом НАДН-цитохром b5 редуктази є ФАД, який бере участь у транспорті електронів з НАДН ⋅ Н+ на цитохром b5 (рис. 23). Наступним акцептором електронів може бути десатураза (цей фермент вводить подвійний зв'язок у структуру жирних кислот) або цитохром Р450 (рис. 21).

Рисунок 22 - Схема електрон-транспортного ланцюга НАДФН-цитохром Р450 редуктазної системи

Рисунок 23 - Схема електрон-транспортного ланцюга НАДН-цитохром b5 редуктазної системи
Цитохром Р450 належить до групи гемопротеїнів, які можуть зв’язувати СО. Саме після зв’язування з монооксидом вуглецю цей цитохром має максимум поглинання світла при довжині хвилі 450 нм і тому отримав назву Р450. Печінка та нирки містять різний ізоферментний спектр цього цитохрому. На сьогодні відомо приблизно 150 генів цитохрому Р450.
Група цитохромів Р450 має низьку субстратну специфічність – вони можуть каталізувати гідроксилювання різноманітних сполук і тому їх називають оксидазами мішаної функції.
Синтез цитохрому Р450 може бути індукований різноманітними сполуками – індукторами. Відомо приблизно 250 хімічних речовин, які індукують синтез ферментів мікросомального окиснення (барбітурати, спирти, кетони, деякі стероїди).
51.
Реакції цього шляху відбуваються під дією активних форм кисню (АФК), які містятьнеспарений електрон і тому є високореакційноздатними – можуть окислювати різноманітні сполуки в організмі і тим самим пошкоджують їх структуру. АФК є агресивними молекулами по відношенню до білків, нуклеїнових кислот і, що саме головне, до ліпідів біомембран. Вони запускають ланцюговий процес перекісного окиснення ліпідів (ПОЛ), що призводить до руйнації мембранних структур.
До АФК належать:
- супероксид-аніон (супероксидний аніон радикал) ( );
);
- перекис водню (Н2О2);
- гідроксильний радикал (ОН•);
- синглетний кисень (1О2);
- NO радикал (NO ∙).
АФК утворюються як бічні продукти роботи дихального ланцюга при неповному відновленні кисню. Повне відновлення кисню супроводжується акцепцією 4 електронів і утворенням ендогенної води. В разі, коли кількість електронів, які транспортуються на кисень, менша, утворюються АФК:

Крім того, ці форми кисню, насамперед супероксидний аніон радикал, можуть утворюватися при функціонуванні деяких ферментів (рис. 24).

Рисунок 24 – Утворення, знешкодження та використання АФК:
СОД – супероксиддисмутаза;
МПО – мієлопероксидаза.
Відомо, що 2-5% кисню, який поглинає організм, перетворюється саме в ці активні молекули, найбільш реакційно здатною з яких є гідроксильний радикал – він може окислити будь-яку молекулу в організмі.
Утворення NO радикала (NO ∙) та подальша його взаємодія з супероксидним аніон радикалом призводять до утворення пероксинітриту (ONOO-), який у незначній кількості може виконувати функцію вазодилятатора, а при високій концентрації викликає ушкодження клітин.

Утворення АФК відбувається постійно в клітинах організму і є звичайним метаболічним процесом. Постійним джерелом АФК є макрофаги, які використовують супероксидний радикал для боротьби із чужорідними організмами (вірусами, бактеріями, злоякісними клітинами). На рис. 24 схематично показано, що супероксидний аніон радикал поступово під дією ферментів СОД та МПО перетворюється у гіпохлорну кислоту, яка є сильним оксидантом і здатна руйнувати чужорідні клітини. Далі гіпохлорна кислота під час реакції з перекисом водню може продукувати синглетний кисень та воду.
Вільнорадикальне окиснення є необхідним для утворення біорегуляторів (простагландинів, лейкотрієнів, тромбоксанів) з арахідонової кислоти.
Лише 2% АФК використовується організмом у процесах, корисних для організму. Решта (98%) АФК має руйнівну дію і може пошкоджувати різні клітинні структури.
Негативний вплив АФК пов'язаний з їх здатністю стимулювати утворення органічних пероксидів (ROOH). Цей процес отримав назву перекисне окиснення.
АФК найбільш активно атакують ненасичені жирні кислоти, що знаходяться у складі фосфоліпідів біомембран. Як було зазначено вище, найбільшу окислювальну здатність має гідроксильний радикал, і саме тому він ініціює ланцюг вільно-радикального окиснення. Гідроксильний радикал забирає водень у СН2-групи поліненасиченої жирної кислоти у складі фосфоліпіду і, таким чином, утворюється ліпідний радикал (L∙).
Розвиток ланцюга відбувається шляхом приєднання кисню:
L • + О2 → LOO •
LOO• + LH → LOOM + LR•.
У результаті утворюються ліпідний пероксирадикал LOO• та пероксид ліпіду LOOH (рис. 24).
Радикали, що утворюються, мають високу реакційну здатність – кожний радикал здатний генерувати утворення декілька нових радикалів. Таким чином, виникає ланцюгова реакція перекисного окиснення ліпідів.
Одним із кінцевих продуктів ПОЛ є малоновий діальдегід:

Визначення концентрації цієї сполуки у крові є важливим показником активності ПОЛ та служить маркером ендогенної інтоксикації при серцево-судинних захворюваннях, діабеті, атеросклерозі тощо.
Ланцюг вільнорадикального окиснення може бути обірваний як за рахунок взаємодії радикалів один з одним, так і в реакціях із компонентами антиоксидантної системи клітини (наприклад, вітаміном Е):
LOO• + L• → LOOH + LH;
L• + вітамін E → LH + вітамін E•;
вітамін E• + L• → LH + вітамін Еокисл.

Рисунок 25 - Механізм розвитку вільнорадикальної реакції ПОЛ
Накопичення АФК та активація перекисного окиснення призводять до низки негативних наслідків у клітині:
1) порушенням рідинно-кристалічної структури ліпопроте-їнів мемебран;
2) зниження міцності та порушення структури мембран клітини, в тому числі мітохондріальної мембрани;
3) структурно-функціональні порушення ферментативних систем дихання та оксидоредуктаз;
4) порушення ферментативних систем синтезу АТФ;
5) ушкодження транспортних мембранних систем;
6) гальмування процесів синтезу білків, нуклеїнових кислот, тощо;
7) руйнація лізосом та вивільнення лізосомальних гідролітичних ферментів;
8) накопичення продуктів неповного окиснення субстратів з можливим подальшим розвитком ацидозу;
9) інактивація деяких коферментів (глутатіону, ліпоєвої кислоти).

