




«акони подразненн€
≤ «акон силових в≥дношень (закон сили)
„им б≥льша сила подразника, тим б≥льша (до певних меж) ≥ б≥олог≥чна реакц≥€.

Ѕ≥олог≥чна
реакц≥€
 а
а
б
в
0 1 2 —ила подразника
–исунок 1.4 Ц «акон силових в≥дношень
0 - 1 Ц допороговий д≥апазон сил.
1 Ц м≥н≥мальна сила подразника, здатна викликати м≥н≥мальну реакц≥ю - пор≥г подразненн€.
ƒл€ збудливих структур характерна така законом≥рн≥сть: чим менший пор≥г подразненн€, тим б≥льша збудлив≥сть структури ≥ навпаки.
1 Ц 2 Ц субмаксимальний д≥апазон сил, в €кому працюЇ закон силових в≥дношень.
2 Ц максимальна сила подразника, здатна спричин€ти найб≥льшу реакц≥ю.
2 - 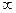 супермаксимальний д≥апазон сил.
супермаксимальний д≥апазон сил.
” цьому д≥апазон≥ сил можлив≥ так≥ вар≥анти в≥дпов≥д≥ б≥олог≥чних структур:
а - збер≥гаЇтьс€ максимальна в≥дпов≥дь;
б - ≥нтенсивн≥сть б≥олог≥чноњ реакц≥њ знижуЇтьс€;
в - б≥олог≥чна структура руйнуЇтьс€, будь-€ка в≥дпов≥дь в≥дсутн€.
≤≤ «акон Увсе або н≥чогоФ
Ќа д≥ю п≥дпорогового подразника б≥олог≥чна структура не в≥дпов≥даЇ (Ун≥чогоФ). Ќа д≥ю подразника пороговоњ сили виникаЇ в≥дразу максимальна в≥дпов≥дь (УвсеФ). ѕодальше зб≥льшенн€ сили подразника не спричин€Ї посиленн€ б≥олог≥чноњ реакц≥њ.
 Ѕ≥олог≥чна
Ѕ≥олог≥чна
реакц≥€
ѕор≥г —ила подразника
ѕодразненн€
–исунок 1.5 Ц «акон Увсе або н≥чогоФ
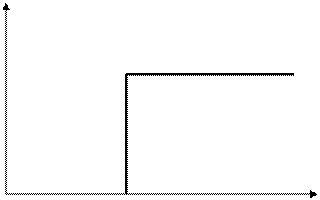
≤≤≤ «акон тривалост≥ подразненн€ (закон Усили часуФ)
„им б≥льша сила подразника, тим менше часу потр≥бно дл€ того, щоб виникла б≥олог≥чна реакц≥€.
–исунок 1. 6 Ц «акон Усили часуФ
2 ‘≤«≤ќЋќ√≤я Ћ≤“»Ќ». ‘”Ќ ÷≤ќЌјЋ№Ќ≤ ¬Ћј—“»¬ќ—“≤ Ћ≤“»ЌЌќѓ ћ≈ћЅ–јЌ»
≈лементарною б≥олог≥чною одиницею Ї кл≥тина. Ќа р≥вн≥ кл≥тини забезпечуЇтьс€ самост≥йне ≥снуванн€ ≥ виконанн€ вс≥х основних б≥олог≥чних функц≥й. Ѕ≥льш≥сть ф≥з≥олог≥чних процес≥в у кл≥тин≥ в≥дбуваютьс€ за участ≥ мембран. ћембрани у кл≥тин≥ виконують так≥ функц≥њ:
Ј —труктурну. —творюють структуру кл≥тини та њњ органоњд≥в.
Ј ≤золюючу. «абезпечують виб≥ркову проникн≥сть кл≥тини до речовин.
Ј —творюють град≥Їнти концентрац≥њ речовин м≥ж в≥дпов≥дними структурами та середовищем, що њх оточуЇ.
Ј –егулюють активн≥сть процес≥в, що в≥дбуваютьс€ у певних структурах ≥ кл≥тин≥ в ц≥лому.
л≥тинна мембрана (плазмалема) Ц це мембрана, що в≥дд≥л€Ї вм≥ст кл≥тини в≥д позакл≥тинноњ р≥дини. ѓњ товщина 7,5 - 10 мкм.
Ѕудова кл≥тинноњ мембрани
—учасною моделлю кл≥тинноњ мембрани Ї р≥динно-мозањчна модель, запропонована у 1972 роц≥ «≥нгером ≥ Ќ≥колсоном (рис.2.1). јвтори модел≥ називали мембрану Ђл≥п≥дним морем, в €кому плавають б≥лков≥ айсбергиї.
ќтже, в≥дпов≥дно до мозањчноњ модел≥ мембрана складаЇтьс€ з таких компонент≥в:
Ј л≥п≥дного компонента (42%);
Ј б≥лкового компонента (55%);
Ј вуглеводного компонента (3%).

–исунок 2.1 Ц –≥динно-мозањчна модель кл≥тинноњ мембрани
Ћ≥п≥дний компонент складаЇ основу мембрани. ¬≥н виконуЇ дв≥ основн≥ функц≥њ:
Ø барТЇрну функц≥ю (розмежуванн€ внутр≥шньокл≥тинного вм≥сту кл≥тини в≥д њњ м≥крооточенн€, транспорт речовин);
|
|
|
Ø матричну функц≥ю (служить матрицею, в €к≥й перебувають численн≥ мембранн≥ б≥лки).
Ћ≥п≥дна пл≥вка €вл€Ї собою подв≥йний шар л≥п≥д≥в, так званий б≥шар, €кий представлений фосфол≥п≥дами ≥ холестеролом.
ћолекула фосфол≥п≥д≥в складаЇтьс€ з г≥дроф≥льноњ гол≥вки ≥ г≥дрофобних хвост≥в (рис.2.2). √ол≥вка становить ¼ молекули фосфол≥п≥ду. ¬она може бути негативно зар€дженою або нейтральною (част≥ше нейтральна, оск≥льки нейтральн≥ гол≥вки легше упаковуютьс€ у пл≥вку, а негативн≥ в≥дштовхуютьс€). ƒо складу гол≥вки вход€ть азотиста основа ≥ фосфорна кислота. ’вости становл€ть ¾ довжини фосфол≥п≥ду. ќдна молекула фосфол≥п≥ду маЇ два хвости. ƒо складу хвост≥в вход€ть вищ≥ жиров≥ кислоти - насичен≥ (пальм≥тинова, стеаринова) ≥ ненасичен≥ (л≥нолева, л≥ноленова, арах≥донова).

–исунок 2.2 Ц —хематичне зображенн€ молекули фосфол≥п≥ду
‘осфол≥п≥дна пл≥вка Ц це р≥дина, €ка дл€ нормального функц≥онуванн€ повинна мати певну вТ€зк≥сть. ” норм≥ вТ€зк≥сть мембрани дор≥внюЇ вТ€зкост≥ оливковоњ ол≥њ. Ќормальна вТ€зк≥сть забезпечуЇтьс€ певним сп≥вв≥дношенн€м насичених ≥ ненасичених жирових кислот: насичен≥ зб≥льшують вТ€зк≥сть, ненасичен≥ зменшують.

–исунок 2.3 Ц ‘ормула фосфол≥п≥ду
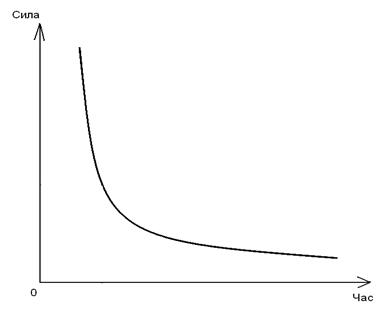
‘осфол≥п≥ди у склад≥ кл≥тинноњ мембрани не закр≥плен≥ жорстко. ¬они рухаютьс€ або у межах одного моношару (латеральна дифуз≥€), або ≥з одного моношару в ≥нший (фл≥п-флоп).
 |
ј Ѕ
–исунок 2.4 Ц ¬иди рух≥в фосфол≥п≥д≥в у мембран≥: ј Ц латеральна дифуз≥€, Ѕ Ц фл≥п-флоп.
’олестерол Ц одноатомний спирт, пох≥дне циклопентанперг≥дрофенантрену. …ого молекула не м≥стить довгих пр€мих ланцюг≥в, а складаЇтьс€ ≥з 4 к≥лець (рис.2.5). ћолекула холестеролу, €к ≥ ≥нш≥ л≥п≥дн≥ молекули маЇ пол€рну ≥ непол€рну частини, тому добре вбудовуЇтьс€ в л≥п≥дн≥ ансамбл≥ кл≥тинних мембран. ѕлазматичн≥ мембрани м≥ст€ть досить значну к≥льк≥сть холестеролу. Ќаприклад, у плазматичних мембранах кл≥тин печ≥нки в≥н становить близько 30% ус≥х мембранних л≥п≥д≥в.
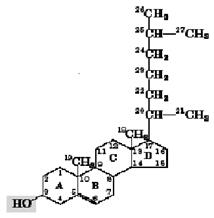
–исунок 2.5 Ц ‘ормула холестеролу
«наченн€ холестеролу дл€ функц≥онуванн€ мембран дуже р≥зноб≥чне.
Ј ’олестерол регулюЇ агрегатний стан б≥л≥п≥дноњ пл≥вки. якщо щ≥льн≥сть кл≥тинноњ мембрани зростаЇ, в≥н розр≥джуЇ њњ. якщо мембрана стаЇ р≥дкою, в≥н, навпаки, робить њњ б≥льш густ≥шою.
Ј ’олестерол Ї мембранним амортизатором. Ћ≥п≥дн≥ ланцюги б≥шару знаход€тьс€ в упор€дкованому стан≥, тому випадков≥ рухи одного з них неминуче передаютьс€ ≥ншим. ’олестерол, вбудовуючись м≥ж фосфол≥п≥дами, блокуЇ цю передачу, тому кооперативн≥ рухи швидко затухають, ≥ в мембран≥ збер≥гаЇтьс€ пор€док.
Ј ’олестерол надаЇ мембран≥ електро≥зол€ц≥йних властивостей.
Ѕ≥лковий компонент. ћолекули мембранних б≥лк≥в наче айсберги плавають у л≥п≥дний матриц≥ мембрани. ѓх под≥л€ють на 2 групи:
1 ≤нтегральн≥ б≥лки Ц це б≥лки, €к≥ проход€ть наскр≥зь мембрани (за х≥м≥чною структурою це переважно гл≥копротењни).
2 ѕериферичн≥ б≥лки Ц це б≥лки, €к≥ не проникають в середину мембрани, а лише прикр≥пл€ютьс€ до њњ внутр≥шньоњ чи зовн≥шньоњ поверхн≥.
‘ункц≥њ мембранних б≥лк≥в:
Ј транспортна. «д≥йсненн€ транспорту речовин через мембрану забезпечуЇтьс€ б≥лками-каналами, б≥лками-переносниками ≥ б≥лками-насосами;
Ј катал≥тична. атал≥з б≥ох≥м≥чних реакц≥й виконуЇтьс€, €к правило, периферичними б≥лками: ендоферментами, €к≥ д≥ють на внутр≥шн≥й поверхн≥ мембрани, ≥ ектоферментами, €к≥ д≥ють на зовн≥шн≥й њњ поверхн≥;
|
|
|
Ј рецепторна. ¬ њњ основ≥ лежить специф≥чна взаЇмод≥€ мембранних б≥лк≥в з р≥зними л≥гандами: мед≥аторами, б≥олог≥чно активними речовинами, гормонами, ≥муноглобул≥нами, компонентами комплементу та ≥н;
Ј антигенна. ѕол€гаЇ у зд≥йсненн≥ ≥мунних реакц≥й;
Ј структурна. Ѕ≥лки забезпечують п≥дтриманн€ певноњ структури кл≥тин. Ќаприклад, спектрин, гл≥кофорин та ≥н.;
Ј зд≥йсненн€ м≥жкл≥тинних взаЇмод≥й забезпечуЇтьс€ адгезивними б≥лками, ≥нтегрином, селектином та ≥н.
¬углеводний компонент кл≥тинноњ мембрани представлений гл≥кокал≥ксом, €кий складаЇтьс€ з вуглеводних залишк≥в мембранних гл≥копротењд≥в ≥ гл≥кол≥п≥д≥в, а також позакл≥тинними протеогл≥канами. “овщина гл≥кокал≥ксу становить приблизно 50 нм. ¬углеводи гл≥кокал≥ксу мають велику к≥льк≥сть ан≥онних груп, що ≥ обумовлюЇ њх основн≥ функц≥њ.
‘ункц≥њ гл≥кокал≥ксу:
Ј створюЇ назовн≥ кл≥тини негативний зар€д. «авд€ки такому зар€ду м≥ж кл≥тинами, що перебувають у р≥дин≥ (наприклад, у кров≥), д≥ють сили в≥дштовхуванн€, ≥ вони не прилипають одна до одноњ;
Ј забезпечуЇ м≥жкл≥тинн≥ взаЇмод≥њ. ” тканинах гл≥кокал≥кс одн≥Їњ кл≥тини може зливатис€ з гл≥кокал≥ксом ≥ншоњ, утворюючи м≥жкл≥тинн≥ контакти;
Ј депонуванн€ позакл≥тинних кат≥он≥в, зокрема, —а2+. «авд€ки пол≥ан≥онному характеру гл≥кокал≥ксу, в≥н може звТ€зувати велик≥ к≥лькост≥ —а2+ ≥, таким чином, служити його депо.
ќсновн≥ в≥дм≥нност≥ х≥м≥чного складу вм≥сту кл≥тини ≥ позакл≥тинноњ р≥дини
≤снують ≥стотн≥ в≥дм≥нност≥ у х≥м≥чному склад≥ внутр≥шньокл≥тинного середовища ≥ позакл≥тинноњ р≥дини.
÷≥ в≥дм≥нност≥ в≥дображено у таблиц≥ 2.1.
“аблиц€ 2.1
| ѕозакл≥тинна р≥дина | ¬нутр≥шньокл≥тинна р≥дина | |
| Na+ | 142 мекв/л | 10 мекв/л |
| K+ | 4 мекв/л | 140 мекв/л |
| Ca2+ | 2,4 мекв/л | 0,0001 мекв/л |
| Mg2+ | 1,2 мекв/л | 58 мекв/л |
| Cl- | 103 мекв/л | 4 мекв/л |
| HCO3- | 28 мекв/л | 10 мекв/л |
| –ќ43- | 4 мекв/л | 75 мекв/л |
| SO42- | 1 мекв/л | 2 мекв/л |
| б≥лки | 5 мекв/л | 40 мекв/л |
| глюкоза | 90 мг% | в≥д 0 до 20 мг% |
| ам≥нокислоти | 30 мг% | 200 мг% |
| рќ2 | 35 мм рт. ст. | 20 мм рт. ст. |
| р—ќ2 | 46 мм рт. ст. | 50 мм рт ст. |
| рЌ | 7,4 | 7,0 |
„ерез кл≥тинну мембрану зд≥йснюЇтьс€ пост≥йний обм≥н речовин м≥ж внутр≥шньокл≥тинним та позакл≥тинним секторами. ќснову цього обм≥ну складають механ≥зми транспорту речовин через кл≥тинну мембрану.
 | |||||||
 |  |  |  |
–исунок 2.6 - ќбм≥н речовин м≥ж кл≥тиною та њњ м≥крооточенн€м
¬езикул€рний транспорт Ц це транспорт речовин за допомогою везикул. ” залежност≥ в≥д напр€мку в≥н буваЇ двох вид≥в:
Ј ендоцитоз (транспорт у кл≥тину);
Ј екзоцитоз (транспорт ≥з кл≥тини).
–озр≥зн€ють два види ендоцитозу:
− фагоцитоз (поглинанн€ твердих речовин);
− п≥ноцитоз (поглинанн€ р≥дких речовин у вигл€д≥ крапель).

–исунок 2.7 Ц ≈ндоцитоз
–ечовина, що поглинаЇтьс€, наближаЇтьс€ до поверхн≥ кл≥тинноњ мембрани ≥ абсорбуЇтьс€ на н≥й. ѕот≥м мембрана вт€гуЇтьс€ всередину ≥ њњ крањ змикаютьс€. ” результат≥ утворюЇтьс€ ендоцитозний пухирець, €кий в≥дриваЇтьс€ в≥д мембрани ≥ м≥груЇ всередину кл≥тини (рис.2.7). Ќайчаст≥ше ендоцитозн≥ пухирц≥ обТЇднуютьс€ в один великий пухирець ≥ зливаютьс€ з л≥зосомою, €ка м≥стить ферменти дл€ перетравленн€ речовин, що транспортуютьс€. ѕродукти г≥дрол≥зу використовуютьс€ кл≥тиною дл€ власних потреб. ѕерш≥ два етапи ендоцитозу в≥дбуваютьс€ без затрат енерг≥њ, тод≥ €к наступн≥ потребують енерг≥њ ј“‘.
Ѕ≥льш≥сть кл≥тин синтезують макромолекули (гормони, б≥лки кров≥, ферменти) "на експорт". р≥м того, у процес≥ обм≥ну речовин в них утворюютьс€ метабол≥ти, €ких треба позбавитис€. ¬иведенн€ цих секрет≥в ≥ екскрет≥в зд≥йснюЇтьс€ за допомогою везикул ≥ називаЇтьс€ екзоцитозом.
–озр≥зн€ють два види екзоцитозу:
− секрец≥€ (вид≥ленн€ ≥з кл≥тини гормон≥в, б≥лк≥в, фермент≥в);
|
|
|
− екскрец≥€ (вид≥ленн€ ≥з кл≥тини продукт≥в обм≥ну).

–исунок 2.8 Ц ≈кзоцитоз
≈кзоцитозн≥ пухирц≥ п≥дход€ть до внутр≥шньоњ поверхн≥ кл≥тинноњ мембрани ≥ контактують з нею за допомогою спец≥альних б≥лк≥в. ќболонка пухирц€ зливаЇтьс€ з мембраною ≥ його вм≥ст опин€Їтьс€ у позакл≥тинному простор≥ (рис.2.8).
“рансмембранний транспорт Ц це транспорт речовини кр≥зь мембрану через вс≥ њњ шари.
“рансмембранний транспорт буваЇ двох вид≥в:
Ј пасивний;
Ј активний.
ѕасивний транспорт Ц це енергонезалежний вид транспорту, €кий зд≥йснюЇтьс€ в≥дпов≥дно до град≥Їнт≥в, що ≥снують в кл≥тин≥.
–озр≥зн€ють так≥ види пасивного транспорту:
Х ф≥льтрац≥ю (транспорт за град≥Їнтом г≥дростатичного тиску)

Х осмос (транспорт за град≥Їнтом осмотичного тиску)

Х дифуз≥ю (транспорт за град≥Їнтом концентрац≥њ, електричних зар€д≥в та ≥н.)

ќсновними видами дифуз≥њ Ї полегшена ≥ проста.
ѕроста дифуз≥€ Ц це дифуз≥€ без використанн€ переносник≥в. ¬она може зд≥йснюватис€ через л≥п≥дний б≥шар ≥ через б≥лки-канали.
„ерез л≥п≥дний б≥шар добре дифундують жиророзчинн≥ сполуки (спирти, кисень, вуглекислий газ, азот) ≥ вода. ¬ода не Ї жиророзчинною сполукою, але вона добре дифундуЇ через л≥п≥дну пл≥вку, оск≥льки њњ молекула маЇ малий розм≥р ≥ високу к≥нетичну енерг≥ю. ≤нтенсивн≥сть дифуз≥њ води через кл≥тинн≥ мембрани дуже велика. Ќаприклад, через мембрану еритроцита кожну секунду дифундуЇ в обидва боки обТЇм води, €кий у 100 раз≥в перевищуЇ обТЇм самого еритроцита. “ранспорт води через л≥п≥дний б≥шар по€снюЇтьс€ Ђтеор≥Їю тимчасових пустотї. «г≥дно з нею через пост≥йний рух хвост≥в фосфол≥п≥д≥в в мембран≥ утворюютьс€ тимчасов≥ пустоти, через €к≥ й проход€ть молекули води.
ѕогано дифундують через л≥п≥дний б≥шар жиронерозчинн≥ сполуки, д≥аметр €ких б≥льший за д≥аметр молекули води, та ≥они. ѕри зб≥льшенн≥ розм≥ру молекули дифуз≥йна здатн≥сть речовини р≥зко зменшуЇтьс€. “ак, д≥аметр молекули глюкози б≥льший за д≥аметр молекули води у 3 рази, а швидк≥сть дифуз≥њ глюкози менша пор≥вн€но з водою у 100 тис€ч раз≥в. ≤они практично не дифундують через л≥п≥дний б≥шар, оск≥льки у водному середовищ≥ мають г≥дратну оболонку, що суттЇво зб≥льшуЇ њх д≥аметр. ƒифуз≥њ запоб≥гаЇ й на€вн≥сть зар€ду ≥он≥в, €кий взаЇмод≥Ї з електричним зар€дом пол€рних головок фосфол≥п≥д≥в.
ƒл€ транспорту ≥он≥в у мембран≥ служать б≥лки-канали (≥нтегральн≥ б≥лки, що вмонтован≥ у мембрану).
Ѕ≥лки-канали мають так≥ властивост≥:
1 —елективн≥сть Ц здатн≥сть виб≥рково пропускати через себе т≥ чи ≥нш≥ сполуки. «алежно в≥д ц≥Їњ характеристики б≥лки-канали бувають неселективн≥ (можуть пропускати р≥зн≥ речовини) ≥ селективн≥ (пропускають переважно один тип молекул). —елективн≥сть каналу може бути абсолютною, коли через канал проходить один тип молекул, ≥ в≥дносною, коли через канал можуть проходити й де€к≥ ≥нш≥ види молекул. —елективними Ї натр≥Їв≥, кал≥Їв≥, кальц≥Їв≥, хлорн≥ та де€к≥ ≥нш≥ канали.
2 Ќа€вн≥сть та стан вор≥тних механ≥зм≥в. ¬орота каналу Ц це структури, в≥д стану €ких залежить стан каналу (ворота зачинен≥ Ц канал закритий, ворота в≥дчинен≥ Ц канал в≥дкритий). ќснову в≥дкриванн€ ≥ закриванн€ каналу складають конформац≥йн≥ зм≥ни б≥лковоњ молекули. ѕричиною цих зм≥н можуть бути 2 типи регул€торних вплив≥в (електричний потенц≥ал та х≥м≥чн≥ речовини). ¬≥дпов≥дно до цього розр≥зн€ють 2 типи контролю стану вор≥т.
1 ѕотенц≥алозалежний механ≥зм. —тан вор≥тного механ≥зму каналу контролюЇтьс€ мембранним електричним потенц≥алом. Ќаприклад, коли мембрана нервового волокна маЇ зар€д -90 м¬, натр≥Їв≥ ≥ кал≥Їв≥ канали закрит≥, коли ж цей зар€д починаЇ зменшуватись, назван≥ канали в≥дкриваютьс€ (рис.2.10).

|
|
|
–исунок 2.10 Ц ѕотенц≥алозалежний механ≥зм контролю стану вор≥т
анали, в €ких стан вор≥т залежить в≥д потенц≥алу на мембран≥, називаютьс€ потенц≥алозалежними
2 Ћ≥гандозалежний механ≥зм. —тан вор≥тного механ≥зму каналу контролюЇтьс€ певними х≥м≥чними сполуками Ц л≥гандами.
ѕри взаЇмод≥њ каналу з х≥м≥чними речовинами зм≥нюЇтьс€ конформац≥€ б≥лка, ≥ канал в≥дкриваЇтьс€. Ќаприклад, при взаЇмод≥њ ацетилхол≥ну з натр≥Ївими каналами мТ€зового волокна ц≥ канали в≥дкриваютьс€ (рис.2.11).
 |

–исунок 2.11 - Ћ≥гандозалежний механ≥зм контролю стану вор≥т
анали, в €ких стан вор≥т залежить в≥д на€вност≥ певних х≥м≥чних речовин, називаютьс€ л≥гандозалежними (хемочутливими).
3 ≥нетика канал≥в, €ка характеризуЇтьс€ швидк≥стю проходженн€ речовин через канал. «алежно в≥д ц≥Їњ характеристики канали под≥л€ють на швидк≥ ≥ пов≥льн≥.
ѕолегшена дифуз≥€ Ц це рух речовин за град≥Їнтом њх концентрац≥њ за участ≥ б≥лк≥в-переносник≥в. ≤нтенсивн≥сть полегшеноњ дифуз≥њ л≥м≥туЇтьс€ к≥льк≥стю молекул б≥лк≥в-переносник≥в та к≥нетикою њх звТ€зуванн€ ≥з речовинами, що перенос€тьс€ (рис.2.12).
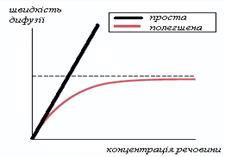
–исунок 2.12 Ц «алежн≥сть ≥нтенсивност≥ полегшеноњ дифуз≥њ в≥д
к≥лькост≥ переносника
≈тапи полегшеноњ дифуз≥њ:
Ј специф≥чне звТ€зуванн€ молекули речовини з б≥лком-переносником;.
Ј конформац≥йн≥ зм≥ни б≥лка-переносника;
Ј внасл≥док цих зм≥н порушуЇтьс€ зв'€зок молекули речовини з м≥сцем њњ звТ€зуванн€ ≥ вона в≥льно переходить на ≥нший б≥к мембрани (рис.2.13).


–исунок 2.13 Ц ћехан≥зм полегшеноњ дифуз≥њ
ћехан≥змом полегшеноњ дифуз≥њ транспортуютьс€ глюкоза ≥ б≥льш≥сть ам≥нокислот.
‘актори, що впливають на ≥нтенсивн≥сть дифуз≥њ речовин через кл≥тинну мембрану
1 ‘актори, повТ€зан≥ ≥з мембраною, через €ку зд≥йснюЇтьс€ дифуз≥€:
а) проникн≥сть мембрани дл€ певноњ речовини - це швидк≥сть дифуз≥њ ц≥Їњ речовини через одиницю площ≥ мембрани на одиницю р≥зниц≥ концентрац≥њ речовини (за в≥дсутност≥ електричного град≥Їнта чи град≥Їнт≥в тиск≥в).
ѕроникн≥сть мембрани залежить в≥д:
Ј товщини мембрани (чим б≥льша товщина, тим менша проникн≥сть);
Ј ф≥зико-х≥м≥чного стану л≥п≥дного шару мембрани. ÷ей стан визначаЇтьс€ х≥м≥чним складом л≥п≥д≥в мембрани: вм≥стом насичених ≥ ненасичених жирових кислот. Ќенасичен≥ жиров≥ кислоти надають мембран≥ р≥дкого стану ≥ зб≥льшують проникн≥сть. ‘≥зико-х≥м≥чний стан мембрани Ї дуже чутливим до температури. ѕри г≥потерм≥њ мембрани Ђтверд≥ютьї ≥ њх проникн≥сть зменшуЇтьс€, а при г≥пертерм≥њ навпаки.
Ј к≥лькост≥ б≥лк≥в-канал≥в та б≥лк≥в-переносник≥в на одиниц≥ площ≥ мембрани та њх функц≥онального стану (закрит≥ вони чи в≥дкрит≥);
б) загальна площа мембрани, через €ку зд≥йснюЇтьс€
дифуз≥€.
2 ‘актори, повТ€зан≥ з властивост€ми ≥ станом речовини, €ка дифундуЇ через мембрану:
а) розчинн≥сть у л≥п≥дах;
б) температура;
в) на€вн≥сть електричного зар€ду;
г) молекул€рна маса.
¬плив молекул€рноњ маси на швидк≥сть дифуз≥њ неоднозначний. « одного боку, чим б≥льша молекул€рна маса речовини, тим б≥льша швидк≥сть руху молекул, тим б≥льша ≥нтенсивн≥сть дифуз≥њ. « ≥ншого боку, зростанн€ д≥аметра молекул ускладнюЇ дифуз≥ю через б≥лки-канали (€кщо д≥аметр молекули б≥льший за д≥аметр каналу, дифуз≥€ через канал припин€Їтьс€).
3 ‘актори, €к≥ Ї руш≥йною силою дифуз≥њ.
≠ град≥Їнт концентрац≥й речовин по обидва боки мембрани;

≤нтенсивн≥сть дифуз≥њ речовини описуЇтьс€ р≥вн€нн€м ‘≥ка
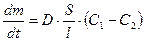 ,
,
де  - ≥нтенсивн≥сть дифуз≥њ;
- ≥нтенсивн≥сть дифуз≥њ;
D Ц коеф≥ц≥Їнт дифуз≥њ;
S Ц площа поверхн≥ мембрани, через €ку зд≥йснюЇтьс€ дифуз≥€;
l - товщина мембрани;
—1-—2 Ц р≥зниц€ концентрац≥й речовини по обидва боки мембрани;
≠ електричний град≥Їнт;

≠ град≥Їнт г≥дростатичних тиск≥в. “ранспорт речовин, €кий зд≥йснюЇтьс€ за град≥Їнтом г≥дростатичного тиску, називаЇтьс€ ф≥льтрац≥Їю. ќск≥льки цей тиск у середин≥ кл≥тини ≥ позакл≥тинному середовищ≥ майже однаковий, то ф≥льтрац≥€ не в≥д≥граЇ ≥стотноњ рол≥ у транспорт≥ речовин через кл≥тинну мембрану, але маЇ суттЇве значенн€, коли йдетьс€ про транспорт через судинну ст≥нку;
≠ град≥Їнт осмотичних тиск≥в. ÷ей град≥Їнт маЇ велике значенн€ дл€ транспорту розчинник≥в, особливо води. “ранспорт за град≥Їнтом осмотичного тиску називаЇтьс€ осмосом.
јктивний транспорт Ц це енергозалежний вид транспорту, €кий зд≥йснюЇтьс€ проти ≥снуючих град≥Їнт≥в (концентрац≥њ, електричного зар€ду, тиск≥в).
«алежно в≥д джерела енерг≥њ, що використовуЇтьс€ на зд≥йсненн€ активного транспорту, його под≥л€ють на первинний ≥ вторинний.
ѕервинний активний транспорт Ц це механ≥зм активного транспорту, що використовуЇ енерг≥ю ј“‘ або ≥нших макроерг≥чних сполук. ¬≥н зд≥йснюЇтьс€ за допомогою б≥лк≥в-насос≥в.
|
|
|
ожен б≥лок-насос складаЇтьс€ з двох компонент≥в:
≠ б≥лка-переносника, що звТ€зуЇ речовину ≥ переносить њњ через мембрану;
≠ б≥лка-фермента Ц ј“‘-ази, що здатна вив≥льнювати енерг≥ю ј“‘ ≥ використовувати њњ дл€ конформац≥йних зм≥н б≥лка-переносника.
ѕрикладами первинного активного транспорту Ї Na+-K+-насос, Ca2+-насос; Ќ+-насос. –озгл€немо принцип роботи насоса на приклад≥ Na+-K+-насоса.
Ќатр≥й-кал≥Ївий насос Ц б≥лок, що зд≥йснюЇ транспорт Na+ ≥з кл≥тини назовн≥ ≥ транспорт K+ у кл≥тину. ÷ей механ≥зм працюЇ в ус≥х кл≥тинах орган≥зму.
Na+-K+- насос складаЇтьс€ з двох субодиниць: великоњ (100 000) ≥ малоњ (55 000). ¬елика субодиниц€ маЇ 3 рецепторн≥ м≥сц€ дл€ звТ€зуванн€ ≥он≥в Na+ на внутр≥шн≥й поверхн≥ мембрани, 2 рецепторн≥ м≥сц€ звТ€зуванн€ ≥он≥в + на зовн≥шн≥й поверхн≥ мембрани ≥ ј“‘-азу на внутр≥шн≥й поверхн≥. як т≥льки в≥дбуваЇтьс€ звТ€зуванн€ 3 ≥он≥в Na+ ≥ 2 ≥он≥в + , внутр≥шн€ частина набуваЇ ј“‘-азноњ активност≥. ≈нерг≥€, €ка вив≥льнюЇтьс€ при розщепленн≥ ј“‘, ≥де на конформац≥йн≥ зм≥ни б≥лка-переносника, ≥ Na+ виноситьс€ з кл≥тини, а + йде у кл≥тину (рис.2.14).

–исунок 2.14 - Ќатр≥й-кал≥Ївий насос
«наченн€ Na+-K+- насоса:
1 «абезпечуЇ р≥зницю концентрац≥й ≥он≥в Na+ ≥ + в кл≥тин≥
≥ позакл≥тинному середовищ≥.
2 —творюЇ електричний потенц≥ал на кл≥тинн≥й мембран≥. ≈лектрогенн≥сть насоса повТ€зана з неекв≥валентним перенесенн€м зар€д≥в п≥д час його роботи (≥з кл≥тини винос€тьс€ 3 позитивн≥ зар€ди (3 Na+), а внос€тьс€ 2 (2 +)). «а один цикл роботи насоса кл≥тина втрачаЇ один позитивний зар€д.
3 ƒотриманн€ сталост≥ обТЇму кл≥тини. як би цей насос не
працював, то б≥льш≥сть кл≥тин набр€кла б.
¬торинний активний транспорт (котранспорт) Ц це механ≥зм активного транспорту, що безпосередньо використовуЇ дл€ транспорту речовин енерг≥ю град≥Їнта концентрац≥њ де€ких ≥он≥в (найчаст≥ше ≥он≥в Na+). ÷ей град≥Їнт, у свою чергу, створюЇтьс€ механ≥змами первинного активного транспорту, а отже, теж залежить в≥д енерг≥њ ј“‘.
¬торинний активний транспорт под≥л€Їтьс€ на два види:
Ј симпорт (транспорт двох речовин в одному напр€мку);
Ј антипорт (транспорт речовин у протилежних напр€мках).
¬торинний активний транспорт зд≥йснюЇтьс€ за допомогою специф≥чних б≥лк≥в-переносник≥в.
—импорт. –озгл€немо механ≥зм симпорту на приклад≥ натр≥Ївого котранспорту глюкози. Ѕ≥лок-переносник, €кий зд≥йснюЇ цей транспорт, маЇ два специф≥чних м≥сц€ звТ€зуванн€ Ц дл€ Na+ ≥ дл€ глюкози. ѕри приЇднанн≥ одного ≥она Na+ ≥ одн≥Їњ молекули глюкози б≥лок зазнаЇ конф≥рмац≥йних зм≥н, у результат≥ чого ≥ Na+, ≥ глюкоза опин€ютьс€ у кл≥тин≥ (рис. 2.15).
«а аналог≥чним механ≥змом в≥дбуваЇтьс€ ≥ натр≥Ївий котранспорт ам≥нокислот. ≤снуЇ 5 тип≥в б≥лк≥в-переносник≥в дл€ транспорту ам≥нокислот. Ќатр≥Ївий котранспорт глюкози ≥ ам≥нокислот особливо ≥нтенсивно в≥дбуваЇтьс€ в еп≥тел≥альних кл≥тинах кишок ≥ тубул€рному еп≥тел≥њ нирок.
р≥м названих механ≥зм≥в котранспорту, у кл≥тинах поширен≥ й ≥нш≥ форми симпорту. Ќаприклад, Na+-K+-Cl- котранспорт, K+-Cl - котранспорт. ” де€ких кл≥тинах ≥снуЇ симпорт ≥он≥в йоду, зал≥за, урат-≥он≥в.

–исунок 2.15 Ц ћехан≥зм симпорту
јнтипорт (≥онообм≥нний механ≥зм). –озгл€немо механ≥зм антипорту на приклад≥ Na+-—а2+-обм≥нного механ≥зму, €кий зд≥йснюЇтьс€ майже в ус≥х кл≥тинах. Ѕ≥лок-переносник, €кий зд≥йснюЇ цей транспорт, маЇ два специф≥чних м≥сц€ звТ€зуванн€ Ц на зовн≥шн≥й поверхн≥ дл€ Na+, на внутр≥шн≥й - дл€ —а2+. ѕри приЇднанн≥ цих ≥он≥в б≥лок зазнаЇ конформац≥йних зм≥н, у результат≥ €ких Na+ опин€Їтьс€ у кл≥тин≥, а —а2+ - поза кл≥тиною (рис. 2.16).

–исунок 2.16 Ц ћехан≥зм антипорту
«а аналог≥чним механ≥змом в≥дбуваЇтьс€ Na+-Ќ+-обм≥нний механ≥зм, €кий зд≥йснюЇтьс€ в еп≥тел≥альних кл≥тинах ниркових канальц≥в, а також Na+-K+, —а2+-Mg2+,Cl--HCO3-, Cl--SO42- - обм≥нн≥ механ≥зми.
3 ѕќ“≈Ќ÷≤јЋ —ѕќ ќё Ќ≈–¬ќ¬»’ ≤
ћТя«ќ¬»’ ¬ќЋќ ќЌ
ћ≥ж внутр≥шньою ≥ зовн≥шньою поверхн€ми плазматичноњ мембрани ус≥х кл≥тин ≥снуЇ р≥зниц€ електричних потенц≥ал≥в. ѓњ називають мембранним потенц≥алом, а в збудливих кл≥тинах Ц потенц≥алом спокою.
ћембранний потенц≥ал (ћѕ) Ц це трансмембранна р≥зниц€ потенц≥ал≥в, €ка ≥снуЇ м≥ж внутр≥шньою та зовн≥шньою поверхн€ми плазматичноњ мембрани.

