




¬ тех случа€х, когда протонефридии уже превратились в метанефри-дии, дело может обсто€ть дво€ко: иногда половые протоки и метанеф-ридии остаютс€ обособленными; у части же полихет происходит сли€≠ние половой воронки с внутренним (отверстым) концом метанефриди€. ¬ результате получаютс€ тоже нефромиксии, но несколько иного свой≠ства, возникающие из сли€ни€ половых воронок не с протонефриди€ми, а с метанефриди€ми (рис. 219, Ѕ; 220, √). Ќередко, особенно у сид€чих Polychaeta, число выделительных каналов сокращаетс€; они сохран€ют≠с€ всего в нескольких сегментах. ќбычно, у сид€чих полихет происходит физиологическа€ дифференциаци€ нефромиксисв на чисто выделитель≠ные и служащие дл€ выведени€ половых продуктов.
роме нефридисв есть другие образовани€, играющие выделительную роль. ¬ некоторых местах, главным образом на стенках кровеносных сосудов, перитонеальный эпителий состоит из крупных клеток, содержа≠щих множество желтых зерен, Ч это накопл€ющиес€ в клетках нераст≠воримые продукты обмена веществ (гуанин или соли мочевой кислоты). Ќаполнившись экскретами, эти, как их называют, хлорагогенные клетки отмирают, а содержимое их поступает в целом и оттуда через нефри≠дии Ч наружу.
Ќервна€ система обнаруживает р€д ступеней усложнени€. “ипична€ центральна€ нервна€ система (см. рис. 217, ј) состоит из парных моз≠говых ганглиев, отход€щих от них и огибающих глотку двух окологло≠точных коннективов и парного брюшного нервного ствола. оннективами называютс€ нервные стволы, соедин€ющие разноименные ганглии (над≠глоточные и подглоточные или ганглии двух соседних сегментов). ѕо≠перечные нервные стволы, св€зывающие ганглии одного сегмента, назы≠ваютс€ комиссурами.
” примитивных форм обе половины брюшного ствола еще широко расставлены и имеют равномерную обкладку из нервных клеток. ƒалее, у части Polychaeta правый и левый брюшные стволы начинают сбли≠жатьс€ к срединной линии; в каждом сегменте на них по€вл€етс€ по скоплению ганглиозных клеток, и оба гангли€ сообщаютс€ поперечной перемычкой (комиссурой) Чполучаетс€ брюшна€ нервна€ система, мет≠ко названна€ нервной лестницей (см. рис. 212). ” большинства полихет стволы сближаютс€ еще больше, оба гангли€ каждого сегмента слива≠ютс€, сами же стволы между гангли€ми либо еще остаютс€ разделенны≠ми, либо тоже сливаютс€, и тогда первичную парность брюшного ствола можно определить только на поперечных разрезах. Ќервна€ система та≠кого устройства называетс€ брюшной нервной цепочкой.
ƒругое постепенное усложнение нервной системы заключаетс€ в ее погружении из кожного эпители€ в полость тела. ¬о врем€ эмбриональ≠ного развити€ нервна€ система закладываетс€ в виде утолщени€ эктодер≠мы. ” некоторых Polychaeta брюшна€ нервна€ система и во взрослом состо€нии остаетс€ в кожном эпителии, не мен€€ своего положени€. ” многих полихет нервные стволы уход€т под кожу и даже под кожно-мускульный мешок, располага€сь в полости тела. ¬ некоторых случа€х, когда р€д сегментов сливаетс€ друг с другом (например, в области глотки), может происходить и соответственна€ концентраци€ нервных узлов.
|
|
|
ќт центральной нервной системы отход€т многочисленные нервы. √оловной мозг посылает от себ€ нервы к антеннам, пальпам и глазам, а каждый ганглий брюшной нервной цепочки иннервирует различные органы соответствующего сегмента.
ќрганы чувств лучше всего развиты у брод€чих полихет. роме эпи≠телиальных чувстительных клеток, рассе€нных в коже, есть специальные
9* 259
органы ос€зани€ и химического чувства. “аковыми служат антенны, пальпы, ресничные €мки, располагающиес€ на простомиуме, и чувстви≠тельные усики параподий.
ќрганы чувства равновеси€ редки, однако у некоторых полихет, осо≠бенно у сид€чих, в передних сегментах тела имеетс€ от 1 до 5 и более пар статоцистов.
ќрганы зрени€ имеютс€ почти у всех многощетинковых. „аще всего глаза расположены на спинной стороне простомиума в числе 2 или 4 и
|
| 8 |
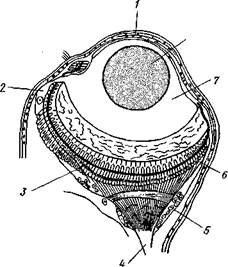
| –ис. 221. √лаз полихеты Alciope (из Ћи≠ванова): / Ч роговица. 2Ч клетка, выдел€юща€ стек≠ловидное тело, 3 Ч светочувствительные па≠лочки, 4 Ч зрительный нерв, 5 Ч зрительный ганглий, 6 ~ сетчатка, 7 Ч стекловидное тело, S Ч хрусталик |
иннервируютс€ от средней части го≠ловного мозга. Ёти надмозговые глаза в наиболее простом случае представл€ют бокаловидное вп€чи-вание эктодермы с суженным отвер≠стием. ¬ыстилающий такую глаз≠ную €мку эпителий играет роль сет≠чатки, а по кра€м вп€чивани€ пере≠ходит в обыкновенный кожный эпи≠телий. —етчатка слагаетс€ из кле≠ток двух сортов. ќдни из них свето≠чувствительны, и они-то, собствен≠но, и €вл€ютс€ насто€щими рети-нальными клетками. ажда€ така€ клетка на конце, обращенном в по≠лость вп€чивани€, т. е. к источнику света, несет светочувствительную палочку, а на другом конце продол≠жаетс€ в нервное волокно. Ќередко, особенно у планктонных форм, на≠пример у Alciope (рис. 221), глаз сильно усложн€етс€: обособл€етс€ от кожи, превраща€сь в замкнутый глазной пузырь, а внутри него диф≠ференцируетс€, кроме дво€ковы≠пуклого хрусталика, еще прозрач≠ное стекловидное тело. ¬се описан≠ные глаза необращенного, или неин≠вертированного, типа. Ќо помимо надмозговых глаз или (при их редук≠ции) вместо них у полихет нередко развиваютс€ глаза на других част€х тела, притом обладающие иным строением. “ак, у многих сид€чих форм, живущих в трубках, развиваютс€ многочисленные глаза на пальпах, превращенных в жабры. ” некоторых мелких форм (јтрџсога и др.), ползающих задним концом вперед, развиваютс€ глазки возле порошицы. Ёти Ђвторичныеї глаза различного строени€.
ѕолова€ система устроена крайне просто. ћногощетинковые коль≠чецы раздельнополы, наружных отличий между полами нет. ѕоловые железы формируютс€ во всех (кроме передних и самых задних) или лишь в некоторых плодущих сегментах под слоем перитонеального эпи≠тели€. „аще всего у основани€ параподий или по соседству с нефриди€-ми половые клетки усиленно дел€тс€ и образуют на стенке целома мест≠ное набухание или половую железу (см. рис. 216; 217, Ѕ). –азвивающие≠с€ гонады сначала прикрыты тонким слоем перитонеального эпители€, который впоследствии лопаетс€, и образующиес€ половые клетки попа≠дают в целом, где они свободно плавают в полостной жидкости и дости≠гают полной зрелости. ” некоторых Polychaeta никаких половых прото≠ков нет, так что живчики или €йца вывод€тс€ наружу просто через раз-
|
рыв стенки тела. ” очень немногих полихет (сем. Capitellidae) имеютс€ самосто€тельные половые воронки с короткими выводными' каналами, открывающимис€ наружу (см. рис. 220, ¬). Ќаконец, в большинстве слу≠чаев эти воронки, как сказано, вступают в сообщение с нефриди€ми, и тогда нефридии несут дво€кую функцию Ч выделение экскретов и выведение наружу половых продуктов. ќплодотворение обык≠новенно наружное.
|
|
|
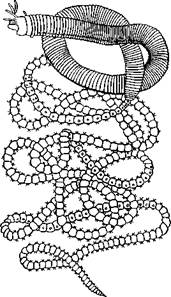
| –ис. 222. “ихоокеанский палоло Eunice viridis (из ‘аусека). ѕе≠редний, более толстый отдел те≠ла, Ч атокный, задний, более тонкий, Ч эпитокный |
Ќередко у полихет в тесной св€зи с по≠ловым размножением находитс€ бесполое, приуроченное к периоду созревани€ поло≠вых продуктов. ѕереходом к этому размно≠жению можно считать так называемые эпи-токные формы многощетинковых. ќбычно наступление половой зрелости не отража≠етс€ па организации черв€. »ногда, однако, плодущие сегменты претерпевают сильную модификацию и в виде так называемой эпигокной части резко отличаютс€ от ос≠тального атокного участка животного. „а≠ще всего эпитокной становитс€ задн€€ по≠ловина черв€. »зменени€ эпитокного участ≠ка сказываютс€ в более сильном развитии параподий и щетинок, в сильной рудимен≠тации кишечника, изменении окраски и т. д. Ёпитокные половозрелые стадии некото≠рых представителей семейств Nereidae, Eunicidae и других поднимаютс€ со дна и при помощи мощных веслообразных пара-подий свободно плавают, приступа€ к поло≠вому размножению. “аков знаменитый па-лоло “ихого океана, который периодиче≠ски в несметном количестве поднимаетс€ со дна и роитс€ в поверхност≠ных сло€х воды, представл€€ в это врем€ лакомую пищу дл€ туземцев (Eunice viridis; рис. 222). ” других видов задн€€ половина тела отрыва≠етс€ от передней, регенерирует на месте разрыва новую голову и, обла≠да€ более мощными параподи€ми, уплывает. Ѕеспола€ половина остает≠с€ на прежнем месте и регенерирует задние сегменты.
” Autolytus (рис. 223, A), Myrianida и некоторых др. эпитокна€ по≠ловина может еще до отделени€ от атокной образовать голову, а беспо≠ла€Ч-начать регенерацию задних сегментов: животное имеет вид двух особей разного строени€, соединенных зоной регенерации. »ногда, нако≠нец, еще до отделени€ первого сформировавшегос€ полового индивида впереди него за счет зоны регенерации образуетс€ втора€, треть€ и т. д. до 30 половых особей, расположенных в один р€д (рис. 223, ј); полу≠чаетс€ временна€ цепочка особей (Autolytus и некоторые другие). «атем половые особи отдел€ютс€ и уплывают (рис. 223, Ѕ, ¬).
–азвитие. ƒробление €йца полное. —начала двум€ последовательны≠ми меридиональными делени€ми €йцо рассекаетс€ на 4 приблизительно одинаковых бластомера: ј, ¬, — и D, причем ¬ отвечает будущей брюш≠ной, a D Ч будущей спинной стороне зародыша. «атем эти 4 клетки де≠л€тс€ наклонной бороздой на 4 мелкие верхние (анимальные) и 4 ниж≠ние крупные (вегетативные) клетки. ѕолучаетс€, как говор€т, квартет микромеров и квартет макромеров (рис. 224). ћикромеры, смотр€ по их происхождению от того или иного макромера, обозначают как la, \b, lc
и Id (рис. 224),макромерыЧ \ј Ч ID. ƒальнейшее дробление состоит в последовательном отделении от макромеров по направлению к ани-мальному полюсу вслед за первым еще 2-го, 3-го и 4-го квартетов мик≠ромеров, при отделении каждого нового квартета клетки ранее образо≠вавшихс€ квартетов тоже дел€тс€.

|
–ис. 223. –азмножение полихет. ј Ч бесполое размножение по-лихеты Autolytus (по √рассе); 5 и ¬ Ч получающиес€ посред≠ством концевого почковани€ по≠ловые особи (5 Ч самка и ¬ Ч самец) тех же червей (по —вешникову)
¬ажна€ особенность отделени€ микромеров заключаетс€ в измене≠нии направлени€ веретена делени€: анимальный конец его отклон€етс€ то по ходу часовой стрелки, то против нее, если наблюдение вести с ани-мального полюса €йца. ≈сли при образовании 1-го квартета €дерные веретена в бластомерах наклонены в одну сторону, то при отделении 2-го квартета все они оказываютс€ отклоненными в другую, и т. д. ¬следст≠вие этого верхние продукты делени€ ложатс€ каждый раз не пр€мо над нижними, а несколько череду€сь с ними, как бы в шахматном пор€дке. Ќапример, микромер \а лежит не целиком над макромером 1Ћ, но отча≠сти налегает и на соседний макромер 1¬. “акое дробление называетс€ спиральным, так как если во врем€ делени€ микромера мысленно про≠должить ось €дерного веретена по сферической поверхности €йца, то по≠лучитс€ спиральна€ лини€.
|
|
|

|
| –ис. 224. —хема спирального дроблени€. ¬ид с верхнего полюса зародыша. ј Ч переход от четы≠рех- к восьмиклеточной стадии; Ѕ Ч стади€ восьми клеток; ¬ Ч переход к стадии 16 клеток; √ Ч за≠родыш на стадии 16 клеток (из «ивинга) |
”же на ранних стади€х дальнейшее развитие каждого эластомера точно определено. ¬ общем, первые 3 квартета микромеров идут на об≠разование эктодермы личинки, один из микромеров 4-го квартета Ч по≠томок клетки D, обозначае≠мый 4d, производит целоми-ческую мезодерму, а осталь≠ные микромеры 4-го кварте≠та и все макромеры дают эн≠тодерму. ¬виду такого ран≠него определени€ дальней≠шего развити€ бластомеров дробление кольчецов назы≠ваетс€ детерминативным. ¬ результате дроблени€ по≠лучаетс€ шаровидна€ бла≠стула, из которой посредст≠вом погружени€ внутрь бла-стоцел€ группы крупных эн-тодермальных клеток про≠исходит гаструла. Ѕласто-пор гаструлы помещаетс€ на ее вегетативном полюсе. «атем бластопор выт€гива≠етс€ по одной из сторон гаструлы (будущей брюш≠ной) к ее экватору, а сзади замыкаетс€. ≈го передн€€ часть превращаетс€ в рото≠вое отверстие. Ќа нижнем полюсе зародыша образует-
с€ порошица. ѕостепенно зародыш превращаетс€ в характерную дл€ полихет личинку Ч трохофору.
“рохофора (рис. 225, ј)Ч типична€ планктонна€ личинка, плаваю≠ща€ при помощи ресничек. “ело ее имеет более или менее шаровидную или эллиптическую форму. Ќа переднем (анимальном) полюсе личинки развиваетс€ чувствительный теменной султан длинных ресниц, сид€щих на группе эктодермальных клеток Ч теменной пластинке. ѕо экватору личинки впереди рта расположен характерный предротовой венчик рес≠ничекЧ прототрох. »ногда позади рта развит менее мощный послеро-товой венчик. ишечник начинаетс€ ртом посредине брюшной стороны личинки и заканчиваетс€ порошицей на ее заднем полюсе и состоит из трех отделов, причем передн€€ и задн€€ кишка образуютс€ вп€чиванием эктодермы, а средн€€ кишка формируетс€ из энтодермы. ћежду кишеч≠ником и стенкой тела находитс€ первична€ полость тела, пересекаема€ тонкими мышечными волокнами. ѕо бокам кишечника лежит пара ма≠леньких протонефридиев.
ћезодермальные органы личинки (главным образом мускульные во≠локна) развиваютс€ из нескольких клеток, лежащих у краев бластопора и называемых мезенхимой. ƒругой мезодермальный зачаток представ≠лен двум€ крупными мезодермальными клетками Ч первичными мезоб-ластами (иногда их называют телобластами)Чпотомками бластомера 4d, лежащими по бокам от кишечника.
ѕосле некоторого периода планктонной жизни трохофоры начинает≠с€ ее метаморфоз. «аднее (вегетативное) полушарие личинки значитель-
но вырастает в длину и подраздел€етс€ сразу на несколько (3, 7, 9Ч13) сегментов. Ќа сегментах развиваютс€ параподии и щетинки или по€в≠л€ютс€ ресничные по€ски. этому времени обе первичные мезодермаль≠ные клетки, усиленно размножа€сь, дают два лежащих по бокам от ки≠шечника клеточных т€жа Ч мезодермальные полоски (рис. 225, Ѕ). ¬ско-

|
| 13 |
–ис. 225. –азвитие Poltjgordius. A Ч трохофора, вид сбоку (по √атчеку); Ѕ Ч метаморфоз трохофоры (по ¬урмбаху):
1 Чанус, 2 Ч задн€€ кишка, 3 Ч средн€€ кишка, 4 Ч мышцы, 5 Ч послеротовой и 6 Ч предротовой венчик ресничек (нрототрох), 7 Ч теменна€ пластинка, 8 Ч те≠менной султан, 9 Ч рот, 10Ч передн€€ кишка, 11 Ч нижнее полушарие трохофоры, 12Ч протонефридий, 13 Чмезодермальна€ полоска, 14 Ч мезобласт, 15 Ч диссепи-
|
|
|
мент, 16 Ч целом
ре под вли€нием наружной сегментации мезодермальные полоски рас≠член€ютс€ на парные группы клеток, так что в каждом сегменте оказы≠ваетс€ сво€ пара мезодермальных клеточных зачатков. ѕоследние сна≠чала компактны, затем в них по€вл€етс€ полость Ч зачаток вторичной полости тела, а ограничивающа€ ее клеточна€ стенка есть стенка цело-мического мешка. ¬ каждом сегменте, таким образом, развиваетс€ пара целомических мешков. ¬ процессе метаморфоза часть клеток теменной пластинки трохофоры погружаетс€ под покровы и образует головной мозг. Ќа брюшной стороне в виде парного валика эктодермы заклады≠ваютс€ брюшные нервные стволы. ¬ дальнейшем они вступают в св€зь с головным мозгом при помощи окологлоточных коннективов. »з экто≠дермы развиваютс€ и органы чувств Ч глаза, пальпы.
“ак, из несегментированной, первичнополостной трохофоры формиру≠етс€ следующа€ личиночна€ стади€ Ч метатрохофора (рис. 226), ха≠рактеризующа€с€ сегментацией и метамерным целомом. “ело сформиро≠ванной метатрохофоры состоит из головной лопасти (простомиума), не≠скольких сегментов и маленькой анальной лопасти (пигидиума) с поро≠шицей на конце. √оловна€ лопасть представл€ет собой почти неизме≠нившеес€ переднее предротовое полушарие трохофоры, а анальна€ ло≠пастьЧ самый нижний участок трохофоры, лежащий позади уровн€ пер≠вичных мезодермальных клеток. “аким образом, оба отдела метатрохо≠форы по природе резко отличны от сегментов туловища. —ущественна€
особенность сегментов метатрохофоры заключаетс€ в том, что все они возникают одновременно.
|
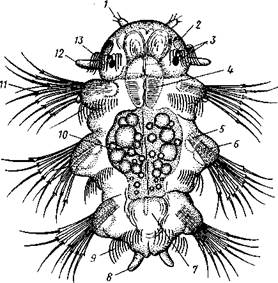
| –ис. 226. ћетатрохофора Nereis pelagica (no –унштрому): / Ч щупальце. 2 Ч пигментна€ клетка. 3Ч глаза, 4 Ч глотка, 5 Ч щетинконосный мешочек параподии, 6 Ч па- раподи€, 7 Ч задн€€ кишка, 8 Ч анальный усик, 9 Ч рес≠ничный по€сок, 10 Ч средн€€ кишка, 11 Чщетинки, 12 Ч пальп, 13 Ч прототрох |
|

|
|

|
| –ис. 227. —хема соотношений между мезодермой лар-вальных и постларвальных сегментов при развитии Polychaeta (по ѕ. »ванову). ј Ч превращение тро-хофоры в мстатрохофору, мезодермальные полоски не сегментированы; Ѕ Ч ћетатрохофора с трем€ лар-вальными сегментами, мезодермальные полоски сег≠ментированы; ¬ Ч по€вление на заднем конце тела эктодермальной зоны роста; √ Ч образование пост≠ларвальных сегментов из зоны роста (зона роста € целомические мешки постларвальных сегментов за≠чернены) |
ћетатрохофора некоторое врем€ плавает или ведет дойный образ жизни, не мен€€сь существенно, а затем испытывает дальнейший мета≠морфоз. Ќа переднем крае анальной лопасти образуетс€ зона роста, клетки которой непрерывно размножаютс€. ќбласть, лежаща€ впереди нее, состоит из быстро растущих, еще не диффе≠ренцированных тканей. ¬ этой зоне формируютс€ новые сегменты и после≠довательно один за дру≠гим отдел€ютс€ по на≠правлению кпереди (рис. 227). ѕроцесс продолжа≠етс€ до тех пор, пока не образуетс€ столько сег≠ментов, сколько их у взрослого черв€. »ногда первичные мезодермаль≠ные клетки сохран€ютс€ в зоне роста и дают нача≠ло мезодермальным поло≠скам. „аше, однако, эти клетки целиком расходу≠ютс€ на образование ие-ломической мезодермы метатрохофоры, и мезо≠дермальные полоски об≠разуютс€ за счет размно≠жени€ эктодермальных клеток зоны роста. ќт ме-зодермальных полосок по≠следовательно отдел€ют≠с€ парные зачатки цело-мических мешков (рис. 227). аждый вновь обра≠зующийс€ сегмент полу≠чает свою пару таких за≠чатков, из которых разви≠ваютс€ его целомические мешки, разрастающиес€ по бокам кишечника квер≠ху и книзу, постепенно вытесн€€ собой первич≠ную полость тела. Ќако≠нец, правый и левый меш≠ки в каждом сегменте встречаютс€ над и под кишкой по спинной и брюшной лини€м, а сле≠дующие друг за другом пары мешков тоже сопри≠касаютс€ своими перед≠ними и задними стенками
(рис. 228). Ќаружным кожно-мускульным листком мешки подстилают эктодерму, а внутренним граничат с кишечником. ¬ результате описан≠ного процесса первична€ полость тела личинки замещаетс€ целомом. «а счет соприкасающихс€ стенок целомических мешков над и под киш≠кой образуетс€ спинна€ и брюшна€ брыжейки, а на месте соприкосно-

–ис. 228. –азвитие целома у кольчатых червей. јЧ¬ Ч поперечные разрезы
трех последовательных стадий развити€ сегмента (из ћатвеева):
/ Ч кишка. 2 Ч первична€ полость тела. 3 Ч целом, 4 Ч наружна€ стенка целомиче-
ского мешка, 5 Ч спинной мезентерий, 6 Ч внутренн€€ стенка целомического мешка,

